Research & Reviews: Journal of Pharmacology and Toxicological Studies
e-ISSN: 2322-0139 p-ISSN: 2322-0120
e-ISSN: 2322-0139 p-ISSN: 2322-0120
Department of Pharmacology, Krishna Institude of Medical Sciences University Karad – 415110, Maharashtra, India.
Received date: 01 December 2012; Revised Date: 28 December 2013; Accepted date: 30 December 2013
Visit for more related articles at Research & Reviews: Journal of Pharmacology and Toxicological Studies
5-hydroxytryptamine (5-HT) inhibits the synthesis and release of dopamine (DA) from rat nigrostriatal neurons. Fluoxetine, a bicyclic antidepressant, enhances serotonergic neurotransmission through potent and selective inhibition of neuronal reuptake of 5-HT. The present study was undertaken to determine whether fluoxetine, through its 5-HT neuronal reuptake inhibiting action, modulates the intensity of the behaviours dependent on the functional status of the nigrostriatal DAergic system. The effect of pretreatment with fluoxetine on dexamphetamine and apomorphine stereotypies of the oral movement variety and on catalepsy induced by haloperidol and small dose of apomorphine was studied in rats. We have also investigated whether fluoxetine induces catalepsy in rats. Our results, that fluoxetine at 2.5, 5, 10 and 20 mg/kg ip neither induced catalepsy nor antagonised apomrphine stereotypy, indicate that fluoxetine at these doses does not block the postsynaptic striatal D2 and D1 DA receptors. It also indicates fluoxetine at 5, 10, 20 mg/kg increase 5-HT neurotransmission by SSRI mechanism and doesnot exert an inhibitory effect at or beyond the postsynaptic striatal D2 and D1, DA receptor sites. Fluoxetine at 5, 10, 20 mg/kg doses increase 5-HT neurotransmission through serotonin SSRI mechanism, inhibit synthesis and release of DA from the nigrostriatal DAergic neurons and thus potentiate catalepsy induced by haloperidol and small dose apomorphine. But, however 10 and 20 mg/kg ip fluoxentine potentiate dexamphetamine stereotypy.
Fluoxetine, apomorphine, dexamphetamine, haloperidol, Stereotypy, Catalepsy, rat.
Ascending serotonergic neurons originating from the midbrain raphe nuclei innervate the substantia nigra pars compacta (SNc), pars reticularis (SNr), ventral tegmental area (VTA), the striatum, nucleus accumbens and the frontal cortex [1,2]. In the SNc and VTA the 5-HTergic fibers make axo-dendritic connections with the dopaminergic cells [1,3]. Autoradiographic and in situ hybridization histochemistry studies have revealed the presence of 5-HT2A and 5-HT2C receptors and their messenger ribonucleic acids (mRNAs) in the SN, VTA, striatum, nucleus accumbens and frontal cortex of rat brain [1,4,5]. A series of in vivo microdialysis and electrophysiological studies have demonstrated that 5-HT2C receptor agonists decrease, whereas 5-HT2C receptor antagonists enhance the release of dopamine (DA) from nigrostriatal, mesolimbic and mesocortical DAergic neurons [6,7,8,9,10]. Based on these observations, 5-HT is considered to exert a tonic inhibitory control on the activity of nigrostriatal, mesolimbic and mesocortical DAergic pathways by stimulating 5-HT2C receptors. On the other hand, 5-HT is proposed to exert a permissive effect on stimulated DA release via activation of 5-HT2A receptors [11] since 5-HT2A agonists enhance 3,4-methylene dioxymethamphetamine induced DA release [12], whereas 5-HT2A receptor antagonists inhibit dexamphetamine induced DA release in striatum and nucleus accumbens [13,14].
Drugs influencing the activity of the central 5-HTergic systems are reported to modulate the intensity of behaviors dependent on the functional status of the nigrostriatal DAergic system, viz DA agonist induced stereotyped behavior [15,16,17] and neuroleptic induced catalepsy [18,19,20,21].
Fluoxetine, a bicyclic antidepressant,enhances serotonergic neurotransmission through potent and selective inhibition of neuronal reuptake of serotonin [22,23,24,25].
Fluoxetine, by blocking the serotonin transporter (SERT), inhibits the reuptake of 5-HT at presynaptic terminals and thus augments the availability of synaptic 5-HT (26).Enhancement of 5-HT synaptic levels in the brain results in activation of various types of 5-HT receptors including the 5-HT2C, 5-HT2A and 5-HT1A receptors [26]. As already stated 5-HT, by activating 5-HT2C and 5-HT2A receptors modulates the activity of the nigrostriatal DAergic neurons [6,13,14,27].
Hyperfunctioning of the nigrostriatal DAergic system in the rat is responsible for the occurrence of stereotyped behavior (SB). The SB manifests as sniffing behavior and of the oral movement variety (OMV) characterized by biting, gnawing and licking behavior. High doses of apomorphine induce sniffing and the OMV of SB by directly stimulating the postsynaptic striatal D2 and D1 DA receptors. The intensity of apomorphine induced SB therefore depends on the functional status of the postsynaptic striatal D2 and D1 DA receptors [28]. High doses of dexamphetamine induce sniffing and OMV of SB by releasing DA from the nigrostriatal DAergic neurons with resultant activation of the postsynaptic striatal D2 and D1 DA receptors by the released DA [28,29]. The intensity of dexamphetamine stereotypy therefore depends on the synthesis of DA and the intraneuronal stores of DA available for release by dexamphetamine, in addition to the functional status of the postsynaptic striatal D2 and D1 DA receptors [28,29].
Hypofunctioning of the nigrostriatal DAergic system in the rat produces a functional lack of DA at the postsynaptic striatal D2 and D1 DA receptors with resultant induction of catalepsy, a state defined as a failure to correct the externally imposed posture by the animal. The neuroleptic haloperidol induces catalepsy by blocking the postsynaptic striatal D2 and D1 DA receptors [30,31]. Small doses of apomorphine selectively stimulate the presynaptic nigrostriatal D2 DA autoreceptors and induce long-lasting inhibition of DA synthesis and release [32,33]. They thus produce a lack of DA at postsynaptic striatal D2 and D1 DA receptor sites with resultant catalepsy in rats [34]. The SB induced by high doses of apomorphine has a rapid onset and is short lasting because of rapid metabolism. In contrast, the small dose apomorphine induced catalepsy has a comparatively delayed onset and is long-lasting [34].
Animals
Albino rats of either sex (weighing 100-180 g), bred in Central Animal House of the Institute, were used. The animals were housed under standard conditions, maintained on a 12 hr light/dark cycle and had free access to food and water up to the time of experimentation. The animals were brought to the department and kept in a noiseless diffusely illuminated laboratory, atleast 1 hr before the experiments for acclimatization to the laboratory environment. Each group consisted of 10 animals. Each animal was used only once. All observations were made between 10 and 17 hrs at 27°-30°C. Observations were made blind with respect to the treatments used. The experimental protocols were approved by the Institutional Animal Ethics Committee and conducted according to the Indian National Science Academy Guidelines for the use and care of experimental animals.
Drugs
Drugs used were fluoxetine hydrochloride (Sun Pharmaceuticals), apomorphine hydrochloride (Sigma), dexamphetamine sulphate (Koch-Light), haloperidol (Senorm injection, Sun), Haloperidol injection solution was diluted to required strength with distilled water. Apomorphine was dissolved in distilled water containing 0.2 mg/ml ascorbic acid while the remaining drugs were dissolved in distilled water only. All drug solutions were prepared immediately before use and were injected intraperitoneally. The volume of injection was 2 ml/kg body weight for the all drugs.
Catalepsy Testing In Rats
For observation and measurement of catalepsy the animals were placed in individual perspex cages (30×20×20cm), 30 min before drug treatment to allow adaptation to the new environment. Animals were tested for catalepsy according to the method of Costall and Naylor [35] by placing both front limbs of the animal over an 8 cm high wooden block and measuring the time that the animal maintained this posture. The animals were considered cataleptic if they maintained this imposed posture for more than 10 sec.
Animals were tested for catalepsy 1.0, 2.0, 3.0 and 4.0 hrs after treatment with fluoxetine (2.5 to 20 mg/kg), haloperidol (1 mg/kg) and distilled water (2 ml/kg body weight ip, control group).
Fluoxetine (FLU) On Stereotyped Behavior (SB) Induced By High Doses of the DA Agonists In Rats
For observation of SB, the rats were placed in individual cages made of wire netting, measuring 30×20×20cm, 30 min before drug treatment to allow adaptation to the new environment.
The intensity of SB was assessed over a 30 sec observation period at 10 min intervals throughout its duration, using the scoring system of Costall and Naylor [35] where periodic sniffing =score 1, continuous sniffing =2, periodic biting, gnawing or licking =3 and continuous biting, gnawing or licking =4. The maximum intensity of SB scored by each rat in the group was taken to compute the mean value of the group.
FLU or haloperidol (HAL) were injected 1 hr before high doses of apomorphine (APO) or dexamphetamine (DAM) treatment. Control groups received distilled water (2 ml/kg body weight ip) 1 hr before receiving APO or DAM.
FLU was tested in the dose range of 2.5 to 20 mg/kg while HAL was tested in the dose of 0.5 mg/kg.
FLU on Catalepsy Induced By HAL and Small Dose of APO in Rats
For observation and measurement of catalepsy the rats were placed in individual perspex cages (30×20×20cm), 30 min before drug treatment to allow adaptation to the new environment. Catalepsy was evaluated by placing both front limbs of the animal over an 8 cm high wooden block and measuring the time for which the animal maintained the imposed posture. Scoring, modified from that of Costall and Naylor, was as follows: maintaining the imposed posture 0-10 sec (0); 11-30 sec (1); 31-60- sec (2); 61-120 sec (3); 121 sec and more (4). Animals were tested and scored for catalepsy 1 and 2 hrs after HAL or small dose APO treatment. Catalepsy score of each animal in the group, at the respective testing time interval, was taken to compute the mean value of the group for that particular timing.
FLU was injected 1 hr before HAL or small dose of APO while the control groups received distilled water (2 ml/kg ip) 1 hr before receiving HAL or small dose of APO.
FLU was tested in the dose range of 2.5 to 20 mg/kg.
In preliminary experiments it was observed that 2.5 to 20 mg/kg FLU produced neither gross behavioral changes nor induced SB of OMV or any feature of the 5-HT1A and 5-HT2A receptor mediated behavioral syndrome in the rats. However, male rats given 5, 10 and 20 mg/kg FLU exhibited the 5-HT2C receptor mediated PEs. As FLU at 40 mg/kg dose had produced ataxia, motor incoordination and muscular hypotonia, for subsequent studies it was therefore used in the dose range of 2.5 to 20 mg/kg.
Induction of Catalepsy In Rats
FLU (2.5, 5, 10 and 20 mg/kg) and distilled water (2 ml/kg body weight ip) treatment did not induce catalepsy in the rats at 1.0, 2.0, 3.0 and 4.0 hrs testing time intervals after administration. Haloperidol (1 mg/kg) however, induced catalepsy in 100% of the animals at all the testing time intervals after administration.
Effect of FLU And HAL Pretreatment on APO Induced SB In Rats
The results are given in Table 1. FLU (2.5 to 20 mg/kg) did not induce SB in rats. APO (1 and 2 mg/kg) induced dose-dependent SB in rats (Table 1).

Table 1: Effect of FLU and HAL pretreatment on APO induced SB in rats.

Figure 1: 
Pretreatment with FLU (2.5 to 20 mg/kg) did not significantly influence APO (1 and 2 mg/kg) induced SB. However, pretreatment with 0.5 mg/kg HAL abolished the SB induced by 1 mg/kg APO and significantly antagonized the SB induced by 2 mg/kg APO (Table 1).
The results are given in Table 2. FLU (2.5 to 20 mg/kg) did not induce SB in rats. DAM (5 and 10 mg/kg) induced dose-dependent SB in rats (Table 2).

Table 2: Effect of FLU and HAL pretreatment on DAM induced SB in rats.
Pretreatment with 2.5 and 5 mg/kg FLU did not significantly affect the intensity of SB induced by 5 and 10 mg/kg DAM. Pretreatment with 10 and 20 mg/kg FLU however, significantly increased the intensity of SB induced by 5 and 10 mg/kg DAM (Table 2). Pretreatment with 0.5 mg/kg HAL abolished the SB induced by 5 and 10 mg/kg DAM (Table 2)

Figure 2: 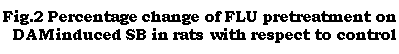
Effect of FLU Pretreatment on HAL And Small Dose APO Induced Catalepsy In Rats
The results are given in Table 3. FLU (2.5 to 20 mg/kg) did not induce catalepsy in rats at both 1 and 2 hr testing time intervals. Pretreatment with 2.5 mg/kg FLU did not significantly affect the cataleptic effect of 1 mg/kg HAL and of 0.1 mg/kg APO at both 1 and 2 hr testing time intervals. Pretreatment with 5, 10 and 20 mg/kg FLU however, significantly increased the cataleptic effect of 1 mg/kg HAL and of 0.1 mg/kg APO at both 1 and 2 hr testing time intervals (Table 3).

Table 3: Effect of FLU pretreatment on HAL and small dose APO induced catalepsy in rats.

Figure 3: 

Figure 4: 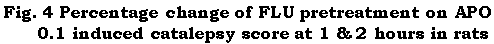
Biochemical and behavioral studies indicate that the midbrain raphe 5-HTergic systems regulate the activity of the nigrostriatal, mesolimbic and mesocortical DAergic systems and modulate the intensity of the behaviors dependent on the functional status of these 3 DAergic systems [26].
In the present study, treatment with 2.5 to 20 mg/kg FLU did not induce OMV of SB in the rats. This indicates that FLU at 2.5 to 20 mg/kg does not stimulate the postsynaptic striatal D2 and D1 DA receptors either directly or indirectly by releasing DA from the nigrostriatal DAergic neurons. Further, treatment with 2.5 to 20 mg/kg FLU did not induce catalepsy, and pretreatment with these doses of FLU failed to antagonize APO-induced SB of OMV in rats. This indicates that at 2.5 to 20 mg/kg FLU does not exert postsynaptic striatal D2 and D1 DA receptor blocking activity. Taken together our findings indicate that FLU does not act as an agonist or antagonist at the postsynaptic striatal D2 and D1 DA receptors. Our results and the conclusion derived thereof concur with the findings in radioligand binding studies that FLU exhibits weak or negligible affinity for the postsynaptic striatal D2 and D1 DA receptors [22,36].
In the present study pretreatment with 2.5 and 5 mg/kg FLU did not have a significant effect on DAM induced SB of the OMV. Pretreatment with 10 and 20 mg/kg FLU however, potentiated DAM induced SB. Since pretreatment with 10 and 20 mg/kg FLU had not potentiated APO induced SB, it suggests that potentiation of DAM stereotypy by 10 and 20 mg/kg FLU is not due to any facilitatory effect of 10 and 20 mg/kg FLU at or beyond the postsynaptic striatal D2 and D1 DA receptor sites.
Potentiation of DAM stereotypy by pretreatment with 10 and 20 mg/kg FLU is explained as pretreatment with 5 to 20 mg/kg FLU is expected to cause accumulation of 5-HT in the synaptic cleft with resultant activation of all the 5-HT receptor subtypes. However, only the 5-HT2A, 5-HT2C, 5-HT3 and 5-HT4 receptors are known to regulate the activity of the nigrostriatal DAergic neurons (26). Furthermore, 5-HT3 and 5-HT4 receptor agonism increases only depolarization evoked or impulse coupled release of DA and not DAM induced or carrier mediated DA release from the nigrostriatal DAergic neurons [26]. Hence activation of the 5-HT3 and 5-HT4 receptors is not responsible for the potentiation of DAM stereotypy by FLU (10 and 20 mg/kg) pretreatment.
5-HT2A receptors are located on the nigrostriatal DAergic neurons. Their activation by 5-HT or 5-HT2A receptor agonists increases DAM induced release of DA from the nigrostriatal DAergic neurons [26]. In contrast, 5-HT2C receptors are located on the striatonigral, striatal and nigral GABAergic neurons. Their activation by 5-HT or 5-HT2C receptor agonists causes release of GABA in the SN and striatum. The released GABA stimulates the GABA B receptors located on the nigrostriatal DAergic neurons and inhibits the activity of the nigrostriatal DAergic neurons [26]. Consequently less DA is available for release by DAM. The influence of FLU pretreatment on DAM stereotypy is thus subject to two opposing effects viz facilitatory via 5-HT2A receptor activation and inhibitory via 5-HT2C receptor activation, the end result depending on which effect predominates.
Pretreatment with 5 mg/kg FLU had exerted 5-HT neuronal reuptake blocking activity. Its pretreatment however, did not significantly influence DAM stereotypy possibly because the two opposing effects might have been of equal magnitude and hence neutralized one another’s effects on the nigrostriatal DAergic neurons. Pretreatment with 10 and 20 mg/kg FLU had potentiated DAM stereotypy possibly because the facilitatory effect of 5-HT2A receptor stimulation on the nigrostriatal DAergic neurons might have predominated resulting in an increase in DAM-induced release of DA and potentiation of DAM stereotypy.
A second possible mechanism by which FLU potentiates the DA-releasing effect of DAM and DAM stereotypy is by inhibiting the metabolism of DAM. DAM is metabolized to the inactive metabolite 4-hydroxyamphetamine by CYP2D1, the rat equivalent of the human CYP2D6 isoenzyme [37]. FLU and norFLU are potent inhibitors of CYP2D [38]. The potentiating effect of FLU pretreatment on amphetamine-induced locomotor activity was due to FLU-induced inhibition of DAM metabolism [39,40]. Even the potentiation of amphetamine activity by the TCA DMI is mainly explained on the basis of its inhibitory action on amphetamine metabolism [41]. Several case reports suggest that patients treated with FLU exhibit an increased sensitivity to DAM though the exact mechanism of interaction between FLU and DAM is not clear [42].
We suggest that pretreatment with 10 and 20 mg/kg FLU potentiates the DA releasing effect of DAM and DAM stereotypy by either or both of the above two stated mechanisms i.e. by activation of 5-HT2A receptors (pharmacodynamic interaction) and/or by inhibiting the metabolism of DAM (pharmacokinetic interaction).
In the present study treatment with 2.5 to 20 mg/kg FLU did not induce catalepsy and pretreatment with these doses of FLU failed to antagonize APO stereotypy. This indicates that FLU at 2.5 to 20 mg/kg does not block the postsynaptic striatal D2 and D1 DA receptors or exerts any inhibitory effect at or beyond the postsynaptic striatal D2 and D1 DA receptor sites.
Potentiation of HAL catalepsy by pretreatment with 5, 10 and 20 mg/kg FLU is explained by us as follows. HAL induces catalepsy by blocking the postsynaptic striatal D2 and D1 DA receptors [31]. In addition, following the blockade of the pre- and postsynaptic nigrostriatal D2 DA receptors by HAL, there is a compensatory ‘feed-back’ increase of nigrostriatal DAergic neuronal activity, which is associated with an allosteric activation of tyrosine hydroxylase. Consequently, there is an increase in the synthesis and release of DA from the nigrostriatal DAergic neurons which counteracts, to some extent, the HAL-induced blockade of the postsynaptic striatal D2 and D1 DA receptors [33].
As already explained earlier pretreatment with 5, 10 and 20 mg/kg FLU exerts 5-HT neuronal reuptake blocking activity and via the accumulated 5-HT in the synaptic cleft causes activation of all the 5-HT receptor subtypes. However, only the 5-HT2A, 5-HT2C, 5-HT3 and 5-HT4 receptors are involved in regulating the activity of the nigrostriatal DAergic neurons [26]. Their role in the potentiating effect of FLU pretreatment on HAL catalepsy is therefore only discussed.
5-HT2A receptor activation increases, whereas 5-HT2C receptor activation decreases, the HAL-induced DA release which occurs during the compensatory ‘feed-back’ increase of the nigrostriatal DAergic neuronal activity due to HAL-induced blockade of the pre- and postsynaptic striatal D2 DA receptors [26]. Consequently 5-HT2A receptor activation, by increasing the release of DA from the nigrostriatal DAergic neurons, will counteract the HAL-induced blockade of the postsynaptic striatal D2 and D1 DA receptors to a greater extent with resultant antagonism of HAL catalepsy. However, 5-HT2C receptor activation, by decreasing the release of DA from the nigrostriatal DAergic neurons, will enhance HAL-induced blockade of the postsynaptic striatal D2 and D1 DA receptors with resultant potentiation of HAL catalepsy. In contrast, 5-HT3 and 5-HT4 receptor agonism has no effect on HAL-induced DA release which occurs during the compensatory ‘feed-back’ increase of nigrostriatal DAergic neuronal activity [26]. Hence activation of the 5-HT3 and 5-HT4 receptors does not have any effect on HAL catalepsy and is not involved in the potentiating effect of FLU (5, 10 and 20 mg/kg) pretreatment on HAL catalepsy.
The end result of FLU pretreatment on HAL catalepsy will therefore depend on whether 5-HT2A or 5-HT2C receptor activation predominates. In our study and that of Khisti et al [43] in rats and of Abdel-Salam and Baiuomy [44] in mice, 5-HT2C receptor activation might have predominated with resultant potentiation of HAL catalepsy. However, in the study of Pires et al [45] in mice 5-HT2A receptor activation might have predominated with resultant antagonism of HAL catalepsy. Even in patients of parkinsonism treated with FLU for associated depression it is possible that only those patients experienced worsening of parkinsonism [46,47,48,49] in whom 5-HT2C receptor activation might have predominated.
HAL is metabolized by CYP2D6 and FLU inhibits CYP2D6 [50,51]. In addition to 5-HT2C receptor activation, a second possible mechanism responsible for or contributing to the potentiating effect of FLU pretreatment on HAL catalepsy might involve FLU-induced inhibition of HAL metabolism [50,51].
Further pretreatment with FLU (5, 10 and 20 mg/kg) potentiated the cataleptic effect of the small dose (0.1 mg/kg) of APO. Potentiation of APO (0.1 mg/kg)-induced catalepsy by FLU (5, 10 and 20 mg/kg) pretreatment is explained as follows. FLU and the small dose of APO reduce the stores of DA in the nigrostriatal DAergic neurons by acting through different mechanisms. FLU, via activation of the 5-HT2C receptors, decreases the nigrostriatal DAergic neuronal activity and synthesis of DA [26]. The small dose (0.1 mg/kg) of APO decreases the synthesis of DA via selective stimulation of the presynaptic nigrostriatal D2 DA autoreceptors [32,33]. Their consecutive administration therefore produces a greater reduction in the intraneuronal stores of DA as compared to the reduction occurring in distilled water pretreated control group receiving 0.1 mg/kg of APO only. A greater reduction in the intraneuronal stores of DA, along with the inhibitory effect of 0.1 mg/kg APO on DA release [33], produces a greater degree of functional lack of DA at the postsynaptic striatal D2 and D1 DA receptor sites. This results in potentiation of the catalepsy induced by the small dose (0.1 mg/kg) of APO.