International Journal of Innovative Research in Science, Engineering and Technology
ISSN ONLINE(2319-8753)PRINT(2347-6710)
ISSN ONLINE(2319-8753)PRINT(2347-6710)
V. Ramamurthy1*, S. Raveendran2, H.V. Anil Kumar3
|
| Related article at Pubmed, Scholar Google |
Visit for more related articles at International Journal of Innovative Research in Science, Engineering and Technology
The aim of this study was to evaluate dietary dosages of Lyngbya majuscula on the immune response and disease resistance against infections due to the opportunistic pathogen Aeromonas hydrophilia, in fish fingerlings. Cyanobacteria, Lyngbya majuscula was incorporated into the diets of fish, Mugil cephalus (Mangrove snapper) fingerlings. Every 20 days, different biochemical, haematological and immunological parameters were analyzed. Superoxide anion production, lysozyme, serum bactericidal, serum protein and albumin were enhanced in cyanobacteria treated groups compared with the control group. After 60 days, fish were survival with Aeromonas hydrophilia, and mortality percentage was recorded up to day 10 post-challenge. Survival decreased in control group (57%) up to day 10 after infection. However, this was increased in the cyanobacteria treatment group, i.e. 85% survivability in the 1 g cyanobacteria/kg, 5 g cyanobacteria/kg and 71% survivability in the 10 g cyanobacteria/kg, respectively. These results indicate that Lyngbya majuscula stimulates the immunity and makes Mugill cephalus more resistant to infection by A. hydrophilia.
Keywords |
| Lyngbya majuscula, Aeromonas hydrophilia, Mugil cephalus, cyanobacteria, Biochemical, Haematological and immunological properties of fish. |
INTRODUCTION |
| The use of immunostimulants in aquaculture is becoming popular for enhancing the activity of non-specific defense mechanisms and increasing disease resistance in mangrove fish. The use of antibiotics and other chemotherapeutics has several drawbacks such as risk of generating resistant pathogens, problems of drug residues accumulating in treated fish and detrimental effect on the environment. Commercial vaccines are expensive for fish producers and may not be available for all species and against emerging diseases1. Therefore, uses of immunostimulants seem to be an alternative way of reducing disease risk in fish culture2. Cyanobacteria are a very old group of organisms and represent relics of the oldest photoautotrophic vegetation in the world that occur in freshwater, marine and terrestrial habitats3. Cyanobacteria have drawn much attention as prospective and rich sources of biologically active constituents and have been identified as one of the most promising groups of organisms to be able of producing bioactive compounds4. Cyanobacteria are known to produce metabolites with diverse biological activities such as antibacterial5-6, antifungal6-7, antiviral8, anticancer9, antiplasmodial10, algicide11, antiplatelet aggregation12 and immunosuppressive2,13 activities. |
| Many bioactive and pharmacologically active substances have been isolated from algae. For instance, extracts of algae were reported to exhibit antibacterial activity opined that the fatty acids (PUFA) in litter fall of mangroves might have positive role on the growth of fishes and shrimps. Screening of cyanobacteria for antibiotics and other pharmacologically active compounds, has received ever-increasing interest as a potential source for new drugs4,14. Cyanobacteria from local habitats seem to be a source of potential new active substances that could contribute to reduction of the number of bacteria, fungi, viruses, protozava and other microorganisms15. Cyanobacteria have not yet been studied for immunostimulants in aquaculture activity and little work has been done to screen cyanobacteria isolated from mangrove with regard to their production of bioactive compounds. The present study was determined the immunostimulating effect of cyanobacteria in mangrove major carp, Mugil cephalus, which is an important species in mangrove water aquaculture. The study was undertaken to evaluate the effects on biochemical, immunological and haematological parameters of the serum/blood of fishes for the first time. |
II. MATERIALS AND METHODS |
| Experimental fish and husbandry: Fingerlings of fish, Mugil cephalus (20 ± 2 g) were collected from, Vedaranyam mangroves, Nagappttinam district, Tamil Nadu. Fishes were stocked in a 50 litre tank and kept for quarantine and health check. After quarantine, fish were acclimatized for 30 days in 40 litre chlorine-free mangrove water and fed with commercial diet. Water exchange (50%) was done daily and water quality was monitored throughout the experiment at three days intervals. Temperature was 28 ± 20C, pH, 7.9 ± 3, salinity 27 ppt, dissolved oxygen concentration 7.0 ± 0.5 mg/l, ammonia-nitrogen concentration 9.8 ± 0.02 mg/l and nitrite-nitrogen 6 ± 0.03 mg/l. Fishes were fed their respective diet at the rate of 4% of body weight per day throughout the experiment. Cyanobacteria: One kilogram of powdered cyanobacteria (Lyngbya majuscula) was collected from the mangrove environs and purified at laboratory in ASN III medium16 and oven-dried at 500C, powdered by mortar and pestle and sieved. For each experiment, the required percentage (1%, 5%, 10% dry weight basis) was included in the feed. These represent diets B, C and D, respectively. Diet A (no cyanobacteria) served as control. Experimental design and feeding diet: Mugil cephalus fingerlings was selected for the study and divided into 4 groups (A, B, C and D). Each group of 10 fingerlings was again divided into two equal duplicate subgroups. Group A was fed with basal diet and acted as the control. The remaining groups were fed with 1 g cyanobacteria/kg of feed (Group B), 5 g cyanobacteria/kg (Group C) and 10 g cyanobacteria/kg of feed (Group D) for experimental periods. Blood and serum samples were collected from fish in each subgroup and examined for the following parameters, total protein, albumin, globulin, albumin/globulin ratio, blood glucose, haemoglobin, serum bactericidal activity, serum lysozyme activity and superoxide anion production, WBC and RBC. Collection of blood: Feed was withheld from fish for 24 hour before blood samples were collected. From randomly picked fish at 20 day intervals, after anaesthetizing with 0.2 ppm MS-222, blood was collected from the caudal vein with a 1 ml plastic syringe ringed with heparin and stored at 40C and used the same day. Blood samples were also collected without heparin, allowed to clot, centrifuged at 3000 rpm for 20 minutes from this supernatant was collected and stored in refrigerated, for estimation of immunological and biochemical parameters. Determination of immunological parameters: The superoxide anion production of blood phagocytes challenged with Aeromonas hydrophilia was measured according to methods of Ramamurthy et al.17. The turbidimetric assay for lysozyme was carried out according to methods of Parry et al.18. Serum bactericidal activity was done following the methods of Kajita et al.19. |
| Determination of blood haematological parameters: Blood haemoglobin content was determined following the cyanomethemoglobin method20. Total erythrocyte count was performed following the method of Hendricks21 using a haemocynometer where a total leucocyte count was determined following the method of Shaw22. Determination of serum/blood biochemical parameters: Serum samples were analyzed for total protein following the method of Lowry et al.23, albumin content by Doumas et al.24, globulin content (subtracting albumin from total protein) and albumin/ globulin ratio. Blood glucose content was estimated following the procedure of Schmidt25. Survival of fish: After 60 days of feeding, 5 fish from each subgroup were challenged intraperitoneally with a lethal dose of Aeromonas hydrophilia and observed for a 20day period for mortality. Biochemical, immunological and enzymatic parameters were assayed in post-challenged groups as per the methods described earlier. |
III. RESULT AND DISCUSSION |
| The result of different dosages of cyanobacteria on production of serum immunological parameters was shown in Table 1. These results indicated that superoxide anion production in the three experimental groups was significantly higher than the control at all the assay periods, except group B in the post-challenge period. Highest superoxide anion production (0.18 ± 0.08) was found in group D fish on day 60. Lysozyme activity in the serum of cyanobacteria fed groups was significantly higher at all sampling times including post-challenge, when compared with the control group. A significant difference was also observed between the treated groups at all sampling times. Highest lysozyme activity (165 ± 0.6) was observed in group C fish on day 60. Serum bactericidal activity in different cyanobacteria fed groups was significantly higher when compared with control at all sampling times, including post-challenge. Highest bactericidal activity (70 ± 0.08) was found in group C fish on day 60. It has been shown that the protective effect of spices may be associated with its antioxidant properties26-27. Neutrophil activity can also be an indicator of the nonspecific response. Cells become more adherent to tissue cell surfaces by the production of adhesion protein, which facilitates their migration from the capillaries to the site of injury28-29. They also exhibit an increased production of oxygen radicals, which are potentially capable of destroying invading pathogens30. In the present experiment, all diets containing various percentages of Lyngbya majuscula showed significant increases in superoxide activity over the control diets, thereby indicating that the level of formation of reactive oxygen species was enhanced. Immunostimulants can increase the non-specific immunity by either increasing the number of phagocytes or activating phagocytosis and respiratory burst31. Post-challenge with Aeromonas hydrophilia showed reduced superoxide activity when compared with the 60-day pre-challenged specimens, but the reduction was not significant. Many things might happen following challenge but it is necessary to speculate that production of superoxide anions by the Lyngbya majuscula in fish act against A. hydrophilia infection. It has been found that Post-challenge with A. hydrophilia showed reduced superoxide activity when compared with the 30-day pre-challenged specimens, but the reduction was not significant1 and aqueous extract of raw garlic and dried powder scavenges hydroxyl radicals32 and superoxide anion33. Similar types of activities might have occurred in the present work. Ramamurthy and Raveendran1 reported that lysozyme activity in the serum of cyanobacteria fed groups was significantly higher at all sampling times including post-challenge, when compared with the control group. A significant difference was also observed between the treated groups at all sampling times. Highest lysozyme activity (145 U/ml) was observed in group III fish on day 30. Immunostimulants can increase serum lysozyme activity, due to either an increase in the number of phagocytes secreting lysozyme or to an increase in the amount of lysozyme synthesized per cell34. Changes in lysozyme activity are greatly influenced by the potency and type of immunostimulants to which fish are exposed. Elevation of lysozyme following immunostimulation has been demonstrated in a number of fish species35. Lysozyme activity was elevated significantly in the groups of fish fed all three levels of cyanobacteria when compared with the control. Serum bactericidal activity was also enhanced in all treated groups when compared with the control group. Many investigators have reported enhanced bactericidal activity by the phagocytic cells of different fish species treated with immunostimulants36. Serum bactericidal activity in different cyanobacteria fed groups was significantly higher when compared with control at all sampling times, including post-challenge2. Blood haematological parameters were shown in Table 1. Haemoglobin content was significantly higher as compared to control only in group D on day 20 and groups C and D on day 40. There was no significant impact of different doses on day 60 and post-challenge. WBC count in different treatments did not show a significant difference on day 20, however, a significantly higher WBC count was found with 5 g cyanobacteria/kg feed and 10 g cyanobacteria/kg feed on day 40 as well as post-challenge and on entire groups B, C and D on day 60. RBC count was significantly higher in all fish fed the different doses of cyanobacteria compared with the control group on all assay days. The number of leucocytes is known to increase sharply when infections occur, as one of the first lines of body defense. The increase in total white blood cell counts and neutrophils, lymphocytes and monocytes counts following 30-day cyanobacteria feeding supports the anti-infection properties of cyanobacyetia37. The erythrocyte count increased with the administration of cyanobacteria, which might indicate an immunostimulant effect and the findings conform to those by Ramamurthy et al.6 who reported that the number of erythrocytes was significantly greater in channel catfish fed with a diet containing b-glucan. The haemoglobin content in the blood and oxygen consumption increases when fishes are under stress. Under such conditions there will be an increase in release of immature RBCs from the haemopoietic organs, which in turn elevate haemoglobin concentration in blood38. In our experiment, the change in haemoglobin content was not significant from control, which indicates the fish was not under stress. |
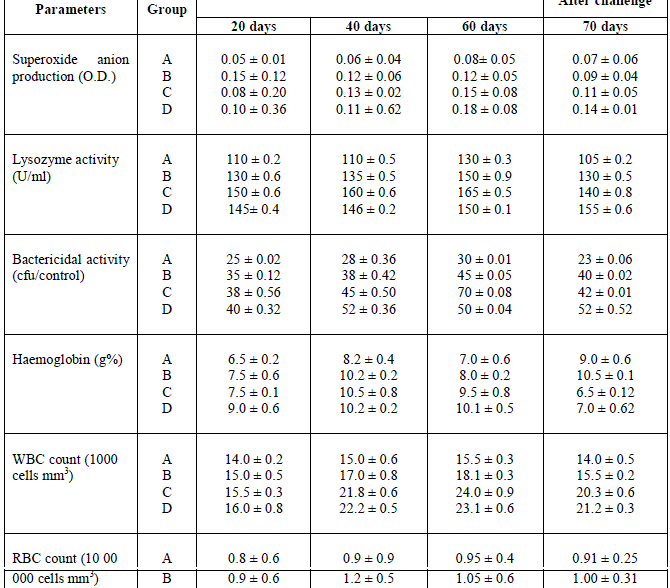 |
| Table 1. Cyanobacteria powder feeding effect on Immunological and Haemtological parameters of Mugil cephalus followed by challenge of Aeromonas hydrophilia after 60 days |
| Serum biochemical parameters are shown in Table 2. Serum protein content was significantly different in groups B and D when compared with group A on day 20. Groups B and C fishes fed for 60 days had significantly higher serum protein content than A group of fish. Post-challenge, significantly higher serum protein content was found in groups C and D when compared with control. Serum albumin content in all fish fed the cyanobacteria doses was significantly higher than the control on day 20, whereas only group D on day 40 and group C on day 60 had significantly higher albumin content than the control fish. Post-challenge, more of the treated fish had elevated serum albumin content compared with control fish. The serum globulin level was significantly higher in group D on day 40 and group C on day 60 in comparison with the control. However, significantly higher globulin content was found in groups C and D when compared with control group A after the post-challenge period. Albumin/globulin ratio was significantly higher in groups B and C fish on day 20, there was no significant elevation in the treated groups thereafter. Blood glucose level at all sampling times was significantly lower in fish fed with different doses of cyanobacteria compared with the control. |
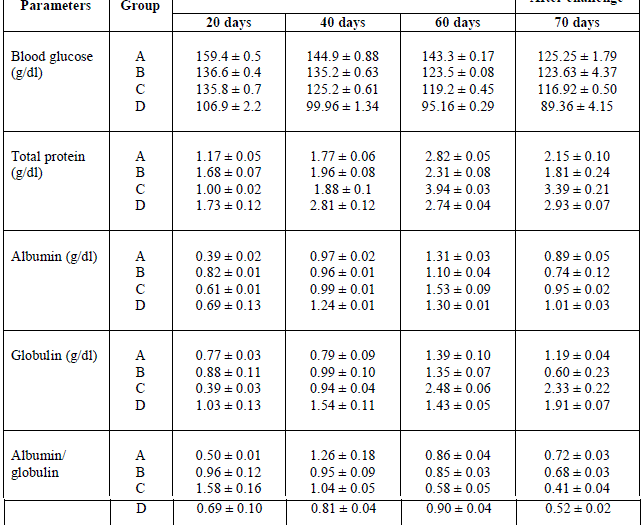 |
| Table 2. Cyanobacteria powder feeding effect on biochemical parameters of Mugil cephalus followed by challenge of Aeromonas hydrophilia after 60 days |
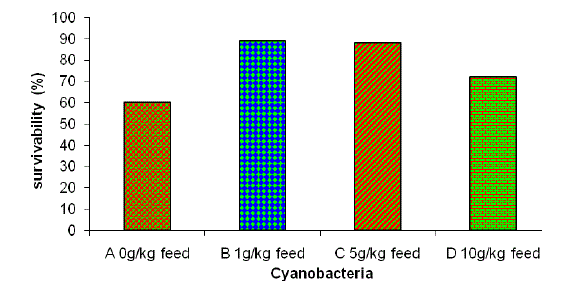
| After challenging fish with Aeromonas hydrophilia, the mortality was recorded for 20 days. There was no mortality of fish up to 12 hours. The group of fish fed with different percentages of cyanobacteria showed higher survival percentage when compared with control. The highest survival (85%) was shown in groups B and C (Fig. 1). The serum total protein after long-term feeding with cyanobacteria increased in comparison to the control diet. According to previously reported1,6 serum protein content was significantly different in groups II and IV when compared with group I on day 10. Groups II and III fishes fed for 30 days had significantly higher serum protein content than group I fish. Post-challenge, significantly higher serum protein content was found in groups III and IV when compared with control. Siwicki39 observed an increase in total protein content after feeding of b-glucan (0.2%) and chitosan (0.5%) in the diet. Serum albumin and globulin values in fish fed with cyanobacteria were higher than the control. Increases in serum protein, albumin and globulin levels are thought to be associated with a stronger innate immune response of fish40. |
 |
| Dietary garlic decreases blood glucose by increasing the level of serum insulin41. According to Sheela and Augusti42, the s-allyl cysteine sulfoxide present in garlic is responsible for its hypoglycaemic activity. Results of the present study indicate that continuous feeding of raw cyanobacterial powder fights against stressors, as was evident from the low glucose value in fishes of groups B, C and D during the experiment. Reduced mortalities against pathogenic challenges at lower dosages of herbal principals were also reported33,43. Citarasu et al.44 developed an Artemia-enriched herbal diet for Penaeus monodon with a combination of five herbs, which significantly increased the growth and survival during stress conditions. Several herbs were tested for their growth-promoting activities in aquatic animals44,45. It is evident from the present work that Lyngbya majuscula could enhance fish immunity after incorporation in feed, even at a lower dose, i.e. 1 g/kg of feed. The present results suggest that inclusion of cyanobacteria in the diet would improve the non-specific immunity of fish and prevent bacterial infections in culture systems. Field trials incorporating these doses merit investigation. Further purification of the active compounds and their evaluation may substantially improve quality as well as their usage in the culture system. |
References |
|