International Journal of Innovative Research in Science, Engineering and Technology
ISSN ONLINE(2319-8753)PRINT(2347-6710)
ISSN ONLINE(2319-8753)PRINT(2347-6710)
| Boitshwarelo Botsheleng1, Thembinkosi Mathowa2, Witness Mojeremane3 UG Student, Department of Crop Science and Production, Botswana College of Agriculture, Gaborone, Botswana 1 Technician, Department of Crop Science and Production, Botswana College of Agriculture, Gaborone Botswana2 Associate Professor, Department of Crop Science and Production, Botswana College of Agriculture, Gaborone Botswana 3 |
| Related article at Pubmed, Scholar Google |
Visit for more related articles at International Journal of Innovative Research in Science, Engineering and Technology
Seed pre-treatments (mechanical, concentrated sulphuric acid for 3, 6, 9 and 12 minutes, hot water treatments for 3, 6, 9 and 12 minutes) were carried out to overcome seed dormancy of Afzelia quanzensis and Baikiaea plurijuga seeds between January and March 2013. Completely randomized designs (CRD) with 4 main treatments (including the control) each replicated 4 times were used. Percentage germination, germination mean time (GMT) and germination index (GRI) were calculated and the data was subjected to analysis of variance (ANOVA). Mechanical scarification significantly (p<0.01) increased the germination of A. quanzensis seeds. Maximum germination percentage (100%) was observed within the shortest GMT of 2.5 days. Also, seed germination was significantly (p<0.01) increased by treating with sulphuric acid for 6 min however, more time was needed to germinate the seeds. All seed pre-treatments except for seeds soaked in sulphuric acid for 6, 9 and 12 min significantly (p<0.01) increased B. plurijuga seeds germination. Maximum germination percentage (100%) was attained with seed soaked in hot water for 9 min. However, hot water treatment revealed superior absolute numbers compared to the rest. Generally, mechanical scarification is the method that suits the two tree species.
Keywords |
| Seeds pre-germination treatments, germination percentage, germination mean time and germination index |
INTRODUCTION |
| Afzelia quanzensis (Welw.) and Baikiaea plurijuga (Harms) are deciduous trees belonging to the family Leguminoseae [1, 2]. A. quanzensis has a wide distribution range in tropical Africa, extending from east African coastal region from Somalia to south Mozambique; widespread in southern Africa, northwards to southern Congo and Angola [1, 2, 3, 4, 5]. It occurs in low-altitude wood land, dry forest and scrub forest [2] up to 1350 m above sea level [5]. The tree grows to a height of 12-15 m, reaching 35 m under ideal conditions [2]. The main truck is short and cylindrical with heavy spreading branches forming a wide crown [3, 6]. The bark is grey-brown, flaking in large pieces leaving pale patches below [2, 3, 6]. The leaves are compound, leafstalk of up to 30 cm with few leaflets [6]. Flowers are usually single petaled and 2.5 to 4.5 cm long, expanded and 2 to 3 cm wide [2, 6]. The timber is used for building, plywood, furniture, paneling etc.[2]. The seeds are used as ornamentals and charms, and are often strung into necklaces or made into trinkets that are sold in curios [2]. The root, bark and leaves of A. quanzensis are used as medicine in many African countries to treat different diseases [2, 5]. |
| B. plurijuga is found in dry deciduous foreston the well-drained Kalahari sands [7]. It occurs in Angola, Botswana, Namibia, Zambia and Zimbabwe [1, 3].B. plurijuga grow to a height of 8-18 m in height, with a trunk up to 60-100 cm in diameter [1] and a large dense and spreading crown[2]. The bark is smooth, later becoming vertically fissured and cracked, and brown to grey in colour [1]. Leaves are alternate, compound, with 4 to 5 pairs of opposite leaflets [2]. The flowers are usually large, attractive, in axial racemes up to 30 cm long; buds are dark brown or golden-brown and densely velvety. Pods are woody, flattened, 13 × 5 cm, broadest near the apex and tapering to the base, with dark brown velvety hairs [2]. Seeds are dispersed when pods split explosively and throwing them some distance [2]. They are dispersed when ground cover is minimal and remain dormant for several months until rain start falling to initiate germination. The wood is heavy, even textured, hard, strong, durable, dark red-brown and slow drying. It is used for furniture, flooring, railway sleepers and mining timber and is exported from southern Africa in considerable quantities [2]. |
| The two species are threatened by increasing rate of exploitation for various uses, wildfires, habitat change and climate change factors. In Botswana, very little effort has been directed towards using indigenous trees in afforestation programmes. The main problem encountered in propagating seedlings of most indigenous trees for afforestation programmes in arid and semi-arid areas is dormant seeds. This dormancy must be broken before germination can occur [8] because it blocks the completion of germination of an intact viable seed under favourable external conditions [8, 9, 10, 11, 12, 13]. Dormancy has evolved differently across species through adaptation to the prevailing environment, to allow seeds to germinate only when conditions are likely to be favourable for a new plant to establish itself [8, 9, 14, 15, 16]. |
| Seed germination starts with the uptake of water by the quiescent dry seed, ending up with the elongation of the embryonic axis [17]. Seeds of most arid and semi-arid tree species areas cannot germinate promptly when subjected to condition favourable for germination due to water impermeable seed coat. Seeds of such species need to be subjected to some physical or chemical treatment to break dormancy and obtain uniform germination. Mechanical or chemical (sulphuric acid) treatment and hot water are the method most commonly used to break water impermeable seed coat to obtain uniform and rapid germination [18, 19, 20, 21, 22, 23, 24, 25, 26]. Little is known about the germination requirements of indigenous trees species in Botswana and therefore the present study was carried to find out how different pre-germination treatments can facilitate germination of A. quanzensis and B. plurijuga seeds. |
II. MATERIALS AND METHODS |
| Two experiments were conducted on A. quanzensis and B. plurijuga in the laboratory at the Department of Crop Science and Production, Botswana College of Agriculture, Botswana from January to March 2013. The Botswana College of Agriculture is on latitude 23°34’S and longitude 25°57’E with an altitude of 994 m above sea level, located at Sebele 10 km from Gaborone along Gaborone to Francistown highway. The seeds were obtained from the National Tree Seed Centre, Ministry of Environment, Wildlife and Tourism, Gaborone, Botswana. The experiments were laid out in completely randomized designs (CRD) with four main (including the control) pregermination treatments each replicated four times. The four pre-germination treatments were as follows; untreated seeds (control), mechanical scarification, chemical scarification and hot water for treatments 1-4 respectively. A total of 800 seeds, 400 seeds for each species were used. The seeds were tested for viability by floating them in distilled water prior to the experiment and those that floated were removed as they may not have been viable whereas, those that sank and settled at the bottom were used in the experiment. Forty (40) glass petri dishes each with a germination paper and allocated 10 seeds were used for each species. Germination papers were kept moistened and distilled water was added whenever necessary throughout the duration of the experiments. Untreated seeds (control) were germinated in petri dishes whereas, mechanical scarification seeds were scarified manually by cutting 1 mm of the seed coat at the opposite site of hilum using a scissor, after which they were germinated in petri dishes. As for chemical scarification, a method described by Emongor et al. [27] was used for this treatment. Seeds were divided and put into four 100 mL heat resistant non-corrosive glass beaker and concentrated sulphuric acid (98%) was poured slowly on the side of the beaker to a level where all the seeds were covered (50 mL). The seeds in the four beakers were left for different times; 3, 6, 9 and 12 min, after which the seeds were removed and the acid drained off into another beaker. Seeds were thoroughly washed and rinsed to remove all the acid in tap water and distilled water, respectively. Seeds were then soaked in distilled water to be ready for germination after 24 hours. Finally, treatment 4 (hot water) seeds were divided and put into four 100 mL heat resistant glass beaker. The seeds in the four beakers were soaked in hot water and left for different times; 3, 6, 9 and 12 min, after which the seeds were removed. Seeds were then soaked in distilled water for 24 hours. |
| The following germination parameters were determined daily; Germination percentage (GP) - the number of germinated seeds as a percentage of the total number of tested seeds given as; |
| GP = (germinated seeds/total tested seeds) × 100 % (1) |
| Germination mean time, was given according toScott et al. [28] as; |
| (GMT days): = ΣTi Ni/S (2) |
| Where Ti is the number of days from the beginning of the experiment, Ni the number of seeds germinated per day and S is the total number of seeds germinated. |
| Germination index (GRI) - was calculated for each treatment using the following equation; |
| GRI = (G1/1) + (G2/2) +......+ (Gx/x), (3) |
| Where G is the germination day 1, 2…, and x represents the corresponding day of germination, [29]. The data collected was subjected to analysis of variance (ANOVA) using the STATISTIX-8 program. Treatment means were separated using Tukey’s Studentized Range (HSD) Test at p≤ 0.05. |
III. RESULTS AND DISCUSION |
| a. Afzeliaquanzensis |
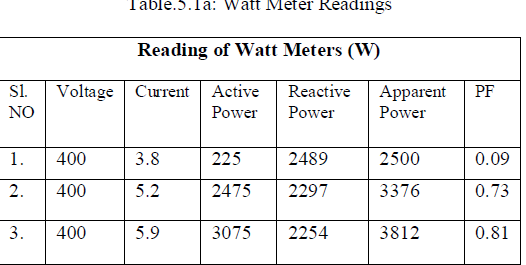
| The germination of A. quanzensis seeds was significantly increased by mechanical scarification (p<0.01) compared to the control after 10 days. Despite differences in absolute numbers, the total means for mechanical scarification, sulphuric acid at 6, 9 and 12 min did not exhibit significant (p>0.05) differences (Table 1). As would be expected mechanical scarification significantly (p<0.01) reduced the germination mean time (GMT) with the lowest of 2.5 days. Moreover, 100% seeds germination was recorded just after 4 days (Table 1). Maximum germination index (GRI) was recorded for mechanical scarified seeds compared to the control which significantly (p<0.01) reduced the seeds GRI (Table 1). |
| The hard seed coat of many arid and semi-arid woody species has evolved to survive under unfavourable conditions such as heat caused by sunlight, severe drought, mechanical and animal damage. Seeds of these species present difficulties to tree growers if sown untreated due to dormancy. Dormancy has evolved differently across species through adaptation to the prevailing environment, to allow seeds to germinate only when conditions are likely to favour the establishment of a new plant [8, 9, 14, 15, 16]. The results of the present study showed that mechanical scarification enhanced the germination of A. quanzensis seeds and significantly reduced the germination mean time. Mechanical scarification is known to break physical dormancy of hard coated seeds which inhibit water uptake and gases such as in Acacia species [30, 31, 32]. Mechanical scarification allows water and air to enter into the seed and stimulate germination. The enhanced germination observed in the mechanical treatment could be attributed to water uptake by the quiescent dry seed, which ended up with the elongation of the embryonic axis [17]. The results of the current study are in agreement with Nainar et al. [33] who used different pre-treatments in Terminalia chebula seeds and found that mechanical scarification gave the highest germination percentage of 60%. The results also showed that immersing A. quanzensis seeds in sulphuric acid for 6, 9 and 12 minutes respectively enhanced seed germination. However, seeds exposed to the same treatment for 12 min declined numerical after 10 days compared to 6 and 9 minutes. This result is supported by several studies which reported that acid treatment is an efficient method of increasing and accelerating seed germination of species with hard impermeable seed coat [34, 35]. Scarifying Lipinus varius seeds in concentrated sulphuric acid was found to increased imbibition, and improved germinationas the duration of scarification time increased [25]. Time of immersing in the acid is critical since long soaking periods can excessively burn the seed coat and damage the embryo [36]. The observed decline in seed germination at 12 min in the present study could be attributed to excessive burning of the seed coat which damaged the embryo. This fact is supported by McDonald et al. [37] who observed that immersing Tamarindus indica and Prosopis africana seedsin sulphuric for more than an hour damage many seeds and significantly reduced germination percentage. Likoswe et al. [35] also reported that soaking seeds of Terminalia sericea for 3-4 hours resulted in no germination. |
| Hot water treatment and the control had low germination percentages not exceeding 40%. This was supported by Centenera et al. [38] that boiling Lupinus hispanicus seed for 240 seconds was not found as a useful treatment compared with mechanical scarification and immersion in sulphuric acid. However, the seeds of A. quanzensis might be heat resistant and probably required longer soaking periods in hot water. Indeed, the results proved the seed dormancy problem as stated by Bewley [8] and the type of dormancy in A. quanzesis is the seed coat dormancy which makes it impossible for water imbibition. |
 |
| b. Baikiaea plurijuga |
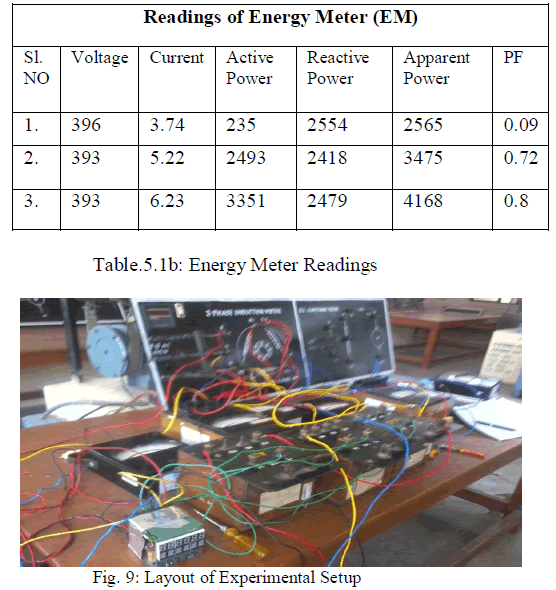
| Mechanical scarified Baikiaea plurijugaseeds had the highest germination percentage (75%) on day 4 followed by sulphuric acid at 3 min with 60%; however, the two treatments were not statistically different whereas the other treatments significantly (p<0.01) reduced the seeds germination percentages (Table 2). Overall, after day 8 hot water treatment across the 3 and 12 min revealed the highest absolute numbers with the highest of 97.5%. However, it was not statistically different from the control, mechanical scarification and sulphuric acid at 3 min (Table 2). The same trend was observed after 10 days where maximum germination of 100% was recorded for hot water at 9 min. The control had a significantly (p<0.01) higher germination mean time (GMT) with the highest of 6.2 days although not statistically different from hot water at 3, 6, 9 and 12 min,whereas sulphuric acid at 12 min significantly (p<0.01) reduced the seeds GMT (Table 2). Maximum germination index (GRI) was recorded in hot water treatment at 9 min however, it was not statistical different from the control, mechanical scarification and hot water at 3, 6 and 12 min (Table 2). |
| Overall, hot water treatment proved to be the best treatment for B. plurijuga seeds as it attained the maximum germination percentage (100%). However, soaking time had a numerical effect on seeds germination although statistically there were no differences. Moreover, the control, mechanical scarification, sulphuric acid at 3 min did not statistically differ from the hot water treatment. This could probably be attributed to the weak seed coat of B. plurijuga seed dormancy and hot water pre-sowing treatment can only be used to accelerate germination. Harper [39] stated that the cause of nature of the seed coat impermeability are not fully understood in some plants, but it has been found that under natural conditions and after most treatments the first site at which water penetrates occurs is the stophioles. This is the weakest and the reinforced area of the seed coat and it is seen as the small raised area close to the hilum but on the side opposite the micropyle. Lower B. plurijuga seeds germination percentages recorded for sulphuric acid could probably be due to the damage caused to the embryo by the chemical [35, 36, 37]. Studies conducted elsewhere showed that sulphuric acid does not improve seed germination of some woody species relative to the control treatment [35, 36]. The result of the present study demonstrate that the germination of B. plurijuga seeds is not limited by the seed coat and hot water treatment can only be used as a way of accelerating germination. |
 |
IV. CONCLUSION |
| Mechanical scarification proved to be very effective in improving seed germination for A. quanzensis species. Also, seed germination was enhanced by treating with sulphuric acid for6 min however; more time is needed to germinate the seeds. Therefore, the type of seed dormancy in A. quanzensis species is physical dormancy. B. plurijuga seeds problem seems not to have an impermeable seed coat since all the treatments performed comparable well except for sulphuric acid treatment at 6, 9 and 12 min which burnt the soft seedcoat and destroyed the embryo. Therefore hot water can only be used to accelerate the germination of B. plurijuga. |
IV. ACKNOWLEDGEMENTS |
| Botsheleng Boitshwarelo was supported by the Ministry of Education Skills and Development scholarship. The authors thank Botswana College of Agriculture for providing resources including laboratory facilities and the National Tree Seed Centre, Ministry of Environment, Wildlife and Tourism for providing seeds. |
References |
|