International Journal of Innovative Research in Science, Engineering and Technology
ISSN ONLINE(2319-8753)PRINT(2347-6710)
ISSN ONLINE(2319-8753)PRINT(2347-6710)
Manish M. Jani1, Jignasa B. Joshi2, Amarnath N. Pandey3
|
| Related article at Pubmed, Scholar Google |
Visit for more related articles at International Journal of Innovative Research in Science, Engineering and Technology
Earth’s surface, due to its heterogeniety, exibits innumerable types of habitates. In general, habitates with thick and fertile soils are characterized as good habitats whereas, degraded lands with thin and nutrient deficient soils are poor habitats. Annul grasses invade and grow abunantly on degraded lands. On the contrary, perennial grasses occupy the habitats with thick and nutrient rich soil [2],[5],[6]. Aristida adscensionsis L. and Melanocenchris jacauemontii J & S. are annul grasses and form a lush green ground vegetation during rainy season on degraded lands in semi arid and arid regions of western India. These annul grasses complete their life cycle extrimely rapidly until moisture is available and persist as dormant seeds during the dry period of the year. Heteropogen contortus L. is a perennial grass and grows on habitats containing relatively thick soil. One form of this species, which is specifically found in western India, lacks rhizome and behaves like annuals. The plants complete their life cycle by the end of monsoon period and therafter dry out. During next monsoon, one to few new tillers appear from the rootstocks which remain alive over long dry period of the year. The present investigation was carried out to understand the adaptation strategies of the aforesaid three grass species to their habitats.
Keywords |
| Heteropogen contortus L, Aristida adscensionsis L., Melanocenchris jacauemontii J & S, Habitat |
INTRODUCTION |
| Grasses and their values have been recognized since time immemorial as the present day cereal crops are the cultivated varieties of their wild ancestors. Use of grasses, as food resources or as fodder has led to extensive breeding programs and improvement in pasture land. In India concept of scientific pasture management has not been properly planned, despite the fact that India has one of the largest livestock populations in the world. Efforts in India for pasture management have been confined either to improvement of existing grasslands or introduction of suitable exotics. There is no sound management plan for the development of pasture land and protection of existing patches of grassland. We have not even fully documented the value of these grasslands in terms of their biological diversity. Among the quarter of a million species of higher terrestrial plants, there is an incredible diversity of form and function. Likewise, there is great variation among habitats in resource availability and abiotic conditions. Plant traits do not vary randomly across ecological gradients and it has long been a central tenet of ecology that environmental conditions are largely responsible for the combinations of traits that have been selected in species and for the sorting of species into assemblages. The repeatable pattern of traits among evolutionarily. In general, habitates with thick and fertile soils are characterized as good habitats whereas, degraded lands with thin and nutrient deficient soils are poor habitats. Annul grasses invade and grow abunantly on degraded lands. On the contrary, perennial grasses occupy the habitats with thick and nutrient rich soil [2],[5],[6]. Aristida adscensionsis L. and Melanocenchris jacauemontii J & S. are annul grasses and form a lush green ground vegetation during rainy season on degraded lands in semi arid and arid regions of western India. These annul grasses complete their life cycle extrimely rapidly until moisture is available and persist as dormant seeds during the dry period of the year. Heteropogen contortus L. is a perennial grass and grows on habitats containing relatively thick soil. One form of this species, which is specifically found in western India, lacks rhizome and behaves like annuals. The plants complete their life cycle by the end of monsoon period and therafter dry out. During next monsoon, one to few new tillers appear from the rootstocks which remain alive over long dry period of the year. The present investigation was carried out to understand the adaptation strategies of the aforesaid three grass species to their habitats. |
MATERIALS AND METHODS |
| Subsoil was filled in the open top cylinders (10 cm diameter X 10 cm depth, cut from PVC pipe) up to varying height of 7.5, 5 and 2.5 cm. The bottom of the cylinders was affixed with wire-net to support soil. The subsoil consisted of mainly pebbles of half decomposed basalt rocks and was collected from 5 – 15 cm depth at the degraded land at Rajkot. Subsequently, fertile black cotton soil was placed above the subsoil and cylinders were completely filled up. Consequently, depth of top soil in the cylinders varied as 2.5, 5.0 and 7.5 cm respectively. Thirty six cylinders were filled up for each depth of top soil. Similar thirty six cylinders were completely filled with black cotton soil, i.e. containing 10 cm thick top soil. Thus, total 144 cylinders were filled with soil containing four depths of top soil. Thereafter, cylinders with each depth of top soil were arranged in four subsets of nine cylinders each. The top soils contained in cylinders of three subsets were separately applied with urea, single superphosphate (P2O5) and mixture of urea and single superphosphate at the rate of 60 kg and 60+30 kgha-1 respectively. Soils contained in remaining nine cylinders of fourth subset for each depth of top soil were not applied with fertilizers and served as control. Subsequently, cylinders for each subset were further arranged in three groups for water treatments. Soils of first group of cylinders were watered at alternate days, those of second group at every fourth days and still those of third group at weekly intervals. Thus soil containing cylinders were arranged in a factorial design with four depths of top soil, four fertilizers treatments and three watering. Sprouting young plants of Hetropogon contortus were dug with intact root system from a protected grassland and soil adhered to underground component of plants was gently washed. The stems above 5 cm from the rootstocks were clipped off. If more than one tillers were present on a rootstock, leaving one, all others were clipped off from their base at rootstock. One rootstock with one 5 cm long stem was transplanted on the top soil of each cylinder. Soil was added tap water to the field capacity (sufficient water to cause drainage). Cylinders were kept in Petri-plates so that leachate caused by watering can be collected and further added to the soil. Thereafter, cylinders were kept inside a glasshouse for 20 days and watered at alternate days. During this period, 2 to 3 leaves emerged on each stem and completed their growth. At this stage plants were considered as fully established. Following established period, plants were given water treatments. |
| Plants were allowed to complete their life cycle and harvested after 35 days since transplantation when about 50% plants turned brown. Plants were washed to remove soil particles adhered with roots. Morphological characters of each plant were recorded. Leaf area was marked on graph sheet. Dry weight of leaves, leaf sheath, stem, inflorescence and root was determined. Root length, which included most of the roots (roughly 95%), was measured. Additional plants grown on 2.5 cm deep top soils, that were watered at alternate days and not treated with fertilizers, were used to determine certain physiological attributes. Leaf area and dry weight of leaves were determined. Water potential (Ψω) of root and leaf was determined by gravimetric method using graded series of solution of sucrose. |
| Collidian solution was applied on the upper and lower leaf surface at midday, dry film were taken off and stomatal number was counted and the size of stomatal aperture was measured under the microscope. Growth study on Aristida adscensionis and Melanocenchris jacquemontii Growth study on Aristida adscensionis and Melanocenchris jacquemontii was conducted following same method that used for the growth study on H. contortus. Apparently identical and young clumps of each species were dug from degraded land at Rajkot. Plants were gently washed to remove soil particles adhered with roots. Tillers were clipped off above 3 cm from their base so that weight of clumps remained almost similar. Clumps were than transplanted on the top soils contained in cylinders. Unexpectedly roots of both the annual species were infected by fungus and plants died within a week. Experiment was repeated but plants did not survive. Following the failure of the experiment, cylinders were filled with sand, as the top soil at degraded land was sandy and nutrient deficient, and plants of Aristida adscensionis and Melanocenchris jacquemontii were transplanted on the top of sands. For each species, nine plants were transplanted separately. Cylinders containing plants were kept inside a glasshouse for 15 days and watered daily. New leaves emerged on stems after 3 to 4 days and plants became established during this period. Following establishment period, plants of each species were given treatments. Cylinders with plants were arranged in three groups. Soil of first group of cylinders were watered at alternate days, those of second group at every fourth day and still those of third group at weekly intervals. Plants were allowed to compete their life cycle and harvested after 25 days since their transplantation when 50% of them turned brown. Plants were washed to separate roots from sands. Morphological characteristics of each plant were recorded using same methods which were followed for the growth study on H. contortus. |
RESULTS AND DISCUSSION |
| Varing depth sof top soil and watering intervals could not alter the leaf area, leaf weight, sheath weight, stem weight and shoot weight of the plants (Table 1- 4). However, urea and single super phosphate promote growth of all the components of plants except for leaf sheath. The effect of urea and single supephosphate fertilizers on the growth of plants was almost similar. The mixture of urea and single superphosphate caused greater influence on the growth of plants than either of fertilizers did separately. Leaf contributed maximum dry matter to the shoot in all the experiments. As a result total shoot weight showed a trend similar to that of leaf under varing conditions. There was an increase in the weight of inflorescence with the application of urea and single superphosphate, but mixture of two fertilizers brought about greater increase than any of them caused separately. Most of the roots were present in 2 to 4 cm depth of top soil and root growth was not influenced by the varing depth of top soil and watering intervals. However, urea and single superphosphate fertilizers separately as well as in mixture promoted the growth of root systems. Root-shoot ratio of plants remained constant under different conditions, including varing depths of top soil, fertilizer treatements and watering intervals. Specific leaf area resaembled to that of xerophytic plants (Table 5). Number of stomata was greater on lower surface than that on upper surface of leaves. Stomatal aperture was narrow. Water potential of leaf and root was high. |
| Growth responce of Aristida adscensionis and Melanocenchris jacquemontii |
| Different components of plants of Aristida adscensionis and Melanocenchris jacquemontii contained very little dry matter (Table 6). It is the characteristic feature of ruderal annual species. Root system was poorly developed and root-shoot ratio was remarkably low. In the spectrum of plant habitats provided by the earth’s surface, the intensities of stress and disturbance vary enormously. Consequently, three major adaptive stretegies have evolved in plants which determine the structure and species composition os a vegetation. These are competitors, which expoit condition of low stress and low disturbance, stress tolerators (high stress low disturbance) and the ruderals (low stress high disturbance) [2]. Considering the natural habitat conditions of low stress and low disturbance at a grassland, H. contortus can easily be placed among copetitors. The term competition [1] as the tendency of neighbouring plants to utilize the same quantum of light, ion of a mineral nutrient, molecule of water. or volume of space. Since H. contortus is a constituent grass species of grassland and completes its life cycle during the monsoon season, most obviously this species can be subjected to competiotio for nutrients. Promotion of growth of the plants of this species by application of urea, single superphosphate and their mixture suggests that nutrient requirement of this species is not saturated by surface soil in natural condition. As a result, nutrient availabilityis the primary factor to limit the occurrence of H. contortus on thick and nutrient rich soils. Roots of the plants penetrated the soil to the depth of 2 to 4 cm suggesting that minimum 2 cm thick surface soil is necessary for the growth of plant of this species. Consequently, thin soil of degraded lands are not suitable for the growth of root system of this species. According to [4] the manner in which a root system develops depends both on internal factors such as its hereditory potentialities and shoot growth depends up on environmental factors such as soil texture, depth, moisture content, aeration, kind and concentration of solutes, and competition with other roots. Resemblance in root-shoot ratio of plants groen under varing conditions of soil depth, fertilizer treatment and watering intervals can be attributed to the relatiion between shoot and root weight. High water potential of root and leaf suggests that plants have not evolved adaptation for drought tolerance. |
| The growth study on Aristida adscensionis and Melanocenchris jacquemontii suggests that aeration was the critical factors for the death of the plants transplanted on soil since plants survived and produced seeds when were grown on sands where aeration is seldom limiting. Inadequate aeration can be further attributed to fungal infection on roots of plants. Attacks on roots by fungi are often increased by deficient aeration because a number of species of pathogenic fungi thrive in poorly aerated soils, and reduced root vigor makes them more susceptible to infection [4], [7], [3]. In general, it appears that oxygen diffusion rates of 0.2 μg cm-2 or less are definitely limiting for most roots of most plants, and rates grater than 0.4 g are adequate, but the adequancy of values in the range between 0.2 and 0.4 depend on factors such as plant species and temperature. However, there is uncertaily conservating the concentration at which carbon dioxide limits growth [4]. Root of these two species spread laterally almost near the top surface of soil. As a result, Aristida adscensionis and Melanocenchris jacquemontii require thin and sandy soils for survival and growth. |
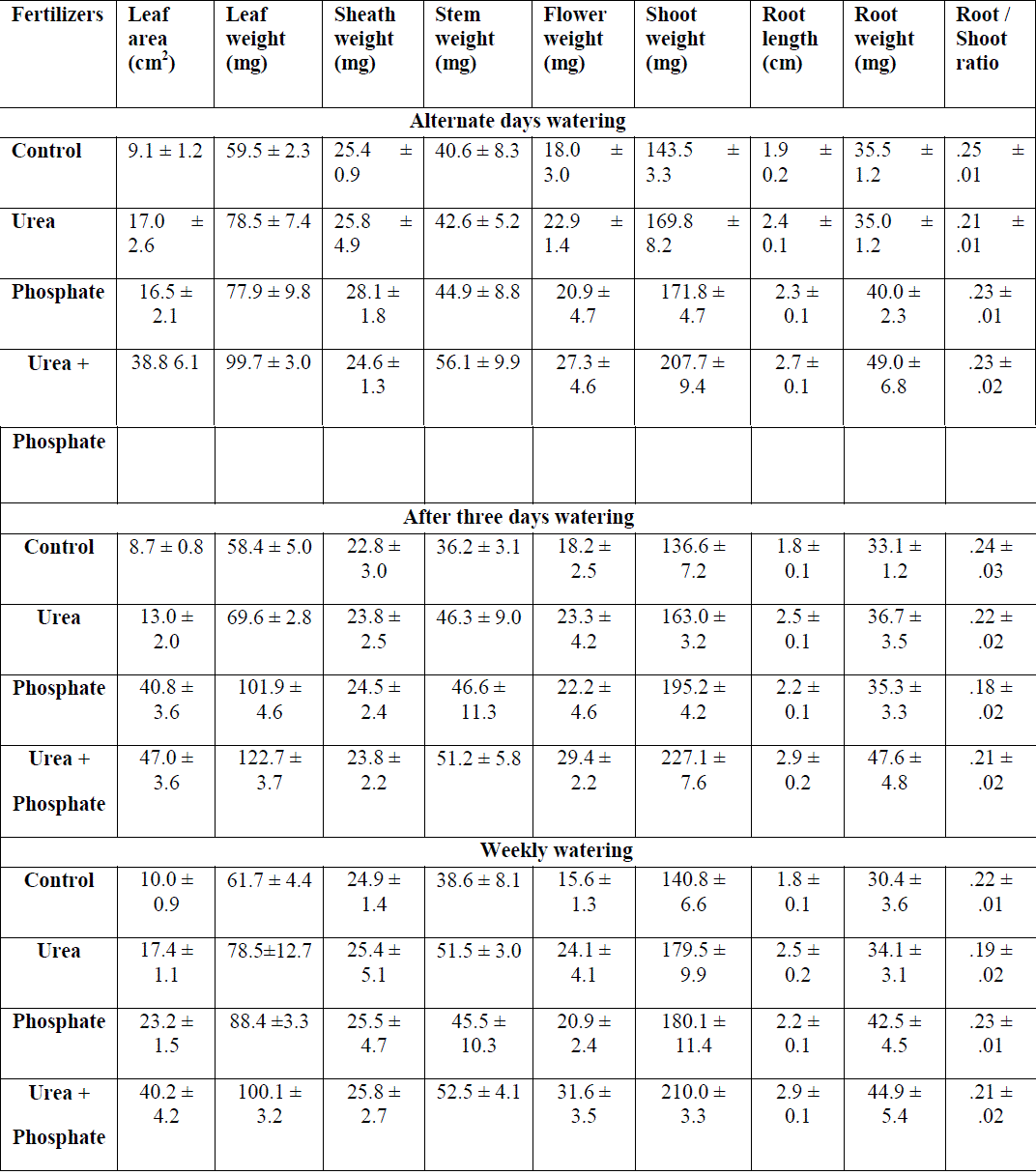 |
| Table: 1 Effect of fertilizers and watering intervals on shoot and root growth of Hetropogon contortus grown on 2.5 cm deep top soil ( ±1 S.E.) |
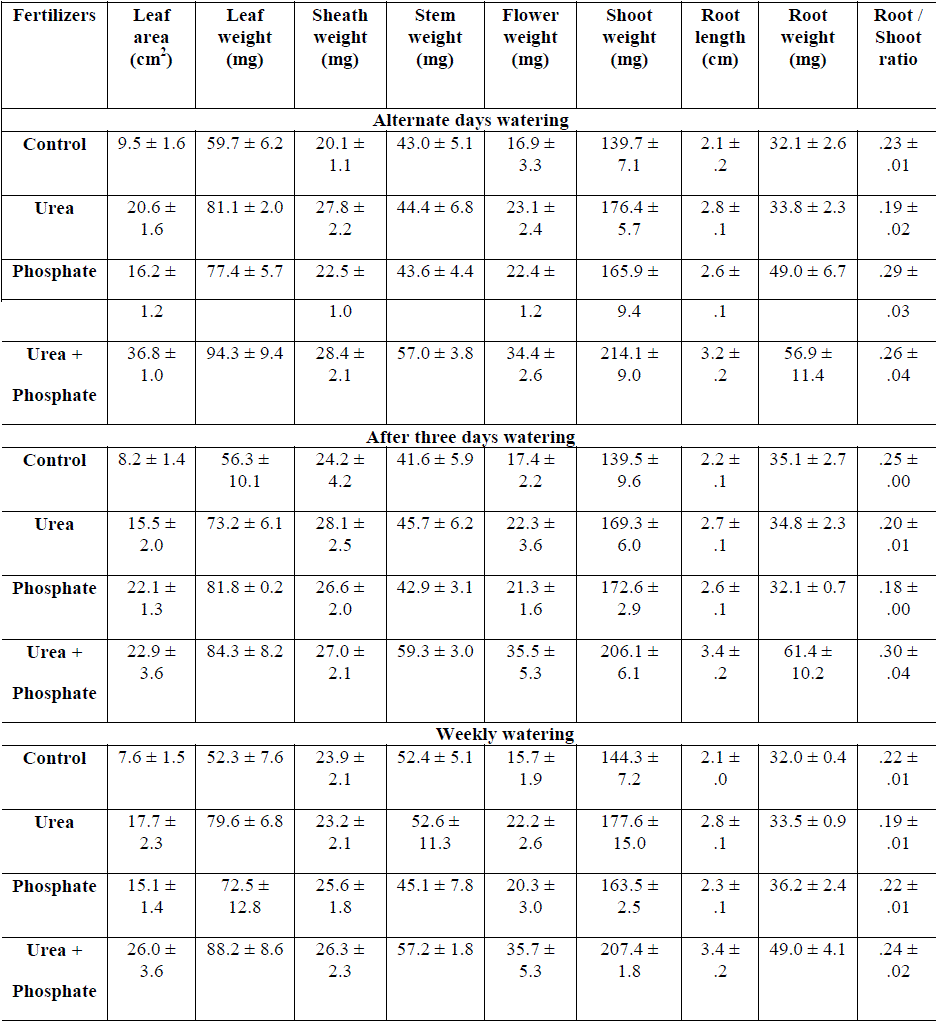 |
| Table: 2 Effect of fertilizers and watering intervals on shoot and root growth of Hetropogon contortus grown on 5 cm deep top soil ( ±1 S.E.) |
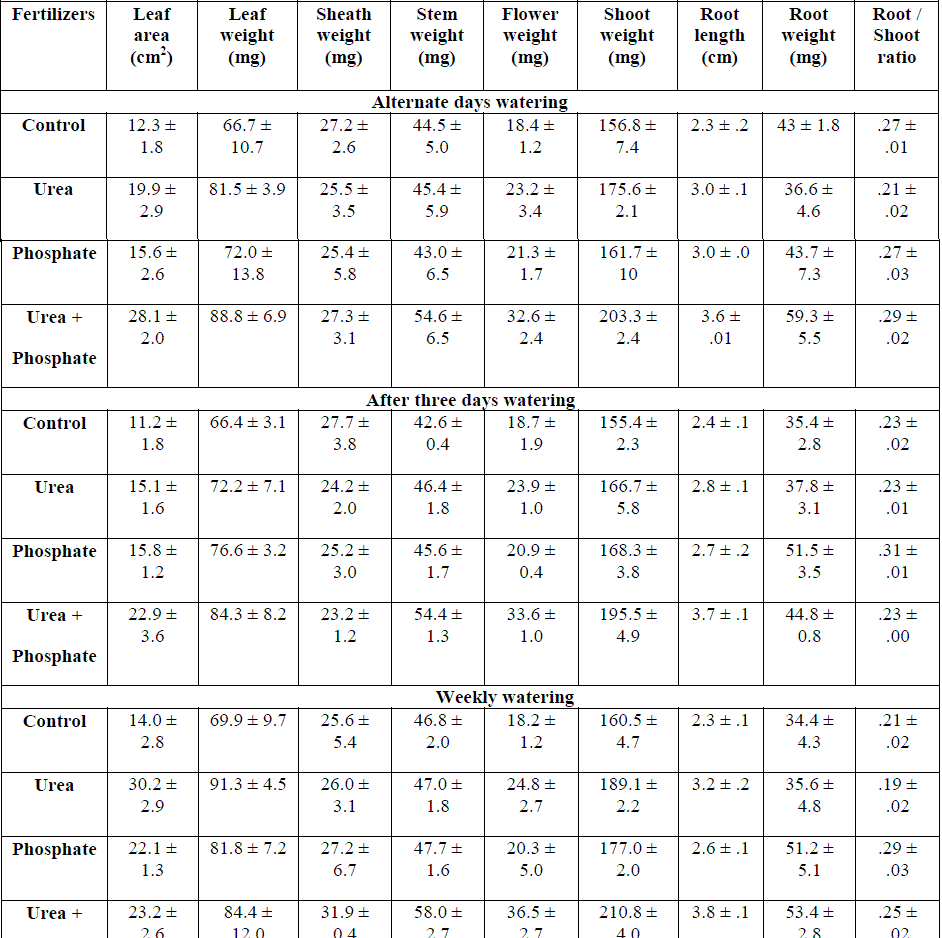 |
| Table: 3 Effect of fertilizers and watering intervals on shoot and root growth of Hetropogon contortus grown on 7.5 cm deep top soil ( ±1 S.E.) |
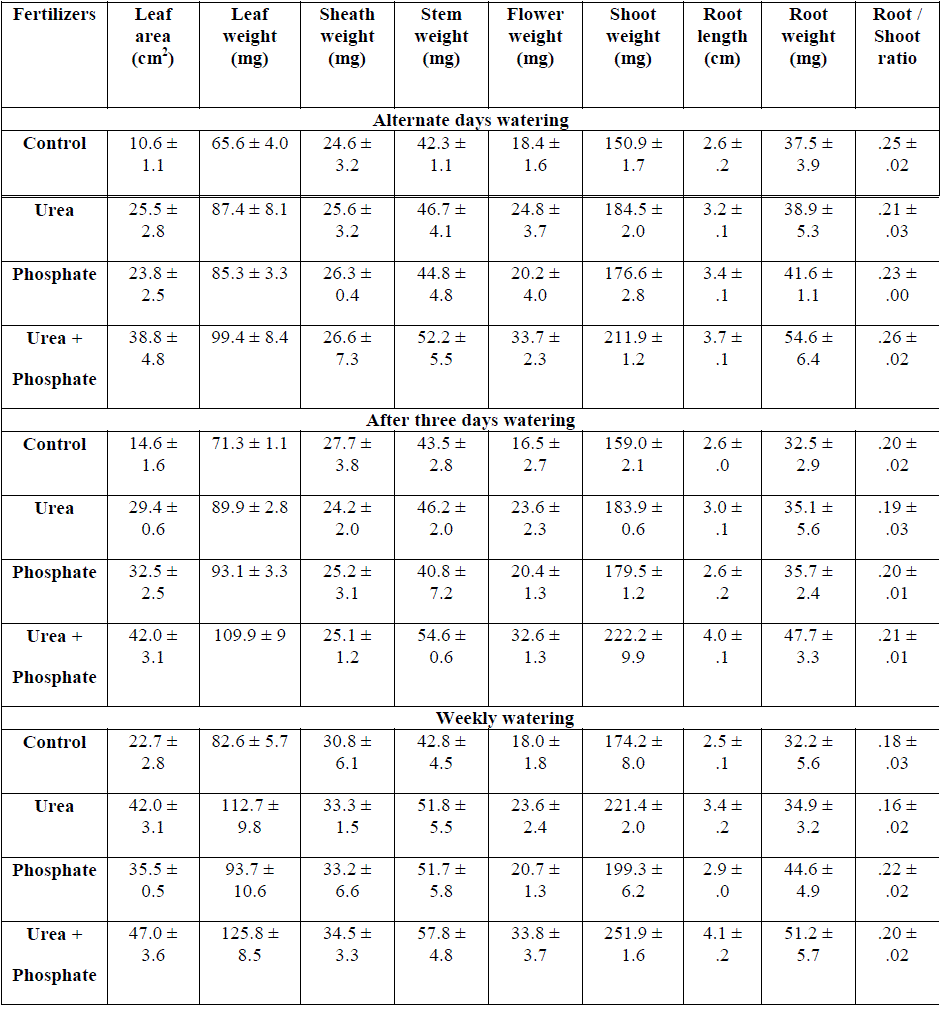 |
| Table: 4 Effect of fertilizers and watering intervals on shoot and root growth of Hetropogon contortus grown on 10 cm deep top soil ( ±1 S.E.) |
 |
| Table: 5 Species leaf area, number of stomata at upper and lower surfaces of leaves, length and width of stomatal aperture and water potential of root and leaf of of Hetropogon contortus ( ±1 S.E.) |
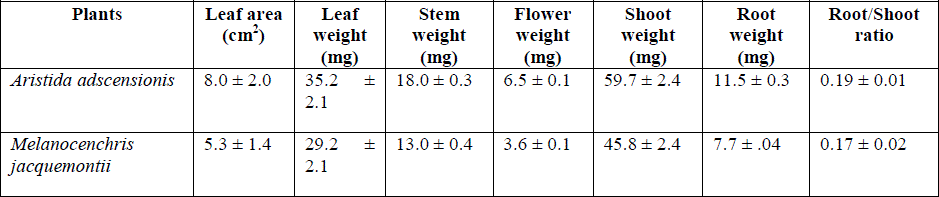 |
| Table: 6 Shoot and root growth of Aristida adscensionis and Melanocenchris jacquemontii on sandy soil containing cylinders (± 1 S.E.) |
CONCLUSION |
| Growth study on Hetropogon contortus Aristida adscensionis and Melanocenchris jacquemontii was conducted to understand their habitat requirement. Nurient availability is major factor to limit the ocuurance of Hetropogon contortus on the habitats with thick and nutrient rich soil. Minimum 2 cm thick soil is necessary for the growth of root system of this species. Consequently this species can not survive on thin and nutrient deficient soil at degraded lands. Aristida adscensionis and Melanocenchris jacquemontii require soil aeration for their survival and susequent growth. As a result, degraded land with thin and sandy soil, where aeration is seldom limiting, are favourable habitats for these two species. |