Research & Reviews: Journal of Zoological Sciences
e-ISSN: 2321-6190 p-ISSN: 2347-2294
e-ISSN: 2321-6190 p-ISSN: 2347-2294
1Department of Biological Sciences, Kyoto University, Oiwake-cho, Kitashirakawa, Sakyo-ku, Kyoto, Japan
2Key Lab of Animal Ecology and Conservation Biology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China
Received date: 11/07/2017 Accepted date: 03/08/2017 Published date: 07/08/2017
Visit for more related articles at Research & Reviews: Journal of Zoological Sciences
While in the evolutionary game theory, the deterministic dynamics is mainly assumed, the stochastic dynamics as well as deterministic dynamics has been assumed in the recent several studies. A previous study developed a stochastic model of two-player games and specified the conditions required for natural selection to favor an invader eventually replacing the population consisting of the resident strategies in finite populations (Nowak et al.). It has been shown that the fixation probability of a single mutant in a population of resident strategies can be greater than 1/N, the fixation probability expected in the absence of natural selection, even when the resident strategy is ESS in the traditional sense. And Nowak et al. has derived the simple rule called the one-third law. Kurokawa and Ihara extended the two player games to n-player games and derived the {2/[n(n+1)]}1/(n–1) law (where n is the group size) as a generalized version of the one-third law. However, the {2/[n(n+1)]}1/(n–1) law holds true for some specific case, and when the law ({2/[n(n+1)]}1/(n–1) law) holds true is unexplored. This paper examines when the law ({2/[n(n+1)]}1/(n–1) law) holds true, and in addition, derives another extension of one-third law (1/[1+(n) 1/(n–1)] law).
Evolutionary game theory, Stochastic process, The one-third law, Two player game, n-player game
In the evolutionary game theory, the deterministic dynamics is mainly assumed [1,2]. However, stochastic dynamics has been focused on since Nowak, et al. [3] was published, and the stochastic dynamics as well as deterministic dynamics has been examined in the recent studies [3-18].
Nowak et al. [3] developed a stochastic model of two-player games and specified the conditions required for natural selection to favor an invader eventually replacing the population consisting of the resident strategies in finite populations. Assuming a discrete-time population of fixed size N with one individual replaced at a time according to the [3,19] Moran model, Nowak et al. showed that the fixation probability of a single mutant in a population of resident strategies can be greater than 1/N, the fixation probability expected in the absence of natural selection even when the resident strategy is ESS in the traditional sense. Furthermore, when both strategy A and strategy B are evolutionarily stable in the traditional sense (i.e., ESS), they demonstrated for sufficiently large population and for sufficiently weak selection that the fixation probability of A is larger than 1/N if p* < 1/3, where p* is the frequency of A at the unstable equilibrium in the conventional deterministic model (i.e., the one-third law).
Lessard, Imhof and Nowak [20,21] calculated the fixation probability on Wright-Fisher processes and showed that one-third law holds true on Wright-Fisher processes as well as on Moran processes. Lessard and Ladret [22] calculated the fixation probability on exchangeable processes including Moran processes and Wright-Fisher processes [23] and showed that one-third law holds true. Ohtsuki, et al. [24] investigated the meaning of one-third law. Hashimoto and Aihara [25] calculated the fixation probability in diploid populations and found ‘‘three-tenth law’’ and ‘‘two-fifth law’’ instead of “one-third law”.
Kurokawa and Ihara, Gokhale and Traulsen [4,8] extended the two player games to n-player games and obtained the fixation probabilities (with which a single mutant eventually replaces the population consisting of the resident strategies). And furthermore, Kurokawa and Ihara [4] have derived the {2/[n(n+1)]}1/(n−1) law (, where n is the group size) as a generalized version of the one-third law.
However, the {2/[n(n + 1)]}1/(n – 1) law holds true for the case where the iterated n-player prisoner’s dilemma is played by a reciprocator, which cooperates only when the every other n−1 group members cooperated in the previous round and an unconditional defector, and payoff is linear. And when the law ({2/[n(n + 1)]}1/(n−1) law) holds true is not explored. In this paper, we examine when the law ({2/[n(n + 1)]}1/(n−1) law) holds true. And furthermore, we derive another extension of one-third law.
Consider the situation where groups of n individuals are formed by randomly choosing individuals from a population, the size of which is N. And a game is played within each of these groups. We consider two strategies A and B, and payoff gained by each individual depends on its own strategy and the strategies of the other n−1 individual in the group. By aj, we denote payoff to an A individual when there are n – j other A individuals in the group, and by bj, we denote payoff to a B individual with n – j A individuals in the group, following [4] (Table 1). The expected payoffs of A and B individuals are described by
| Strategy of the Focal Individual | Number of A Individuals among the n-1 Opponent Players | |||||
| n-1 | n-2 | n-3 | … | 1 | 0 | |
| A | a1 | a2 | a3 | … | an-1 | an |
| B | b1 | b2 | b3 | … | bn-1 | bn |
Table 1: The payoff matrix of the n-player game.
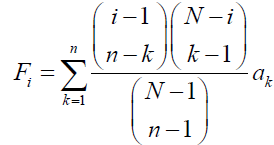 (1)
(1)
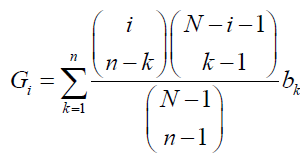 (2)
(2)
Respectively, where i is the number of A individuals in the population (0 ≤ i ≤ N). Note that when l ≥ m is met, 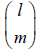 represents a binomial coefficient and otherwise is defined as 0. The fitness of A and B individuals when there are i A individuals
in the population are respectively given by
represents a binomial coefficient and otherwise is defined as 0. The fitness of A and B individuals when there are i A individuals
in the population are respectively given by
fi = 1 − w + wFi (3)
gi = 1 − w + wGi, (4)
where w is the selection intensity and specifies the contribution of the game to fitness [3]. Here, it is assumed that selection intensity is sufficiently weak (0 < w << 1). Population dynamics are formulated as a Moran process with frequency-dependent fitness. Concretely, at each time step, an individual is chosen for reproduction with the probability proportional to its fitness. And one identical offspring is produced to replace another individual randomly chosen for death with the probability 1/N [3,16]. In this process, there are two absorbing states, i = 0 and i = N. Kurokawa & Ihara and Gokhale & Traulsen [4,8] have shown, by using (1)- (4), that in the limit of large population size, the condition under which the fixation probability of A, ρA, with which a population at state i = 1 eventually reaches state i = N is over 1/N is given by
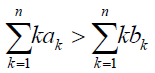 (5)
(5)
Similarly, by using (1)-(4), the condition under which the fixation probability of B, ρB, with which a population at state i = N − 1 eventually reaches state i = 0 is over 1/N is given by
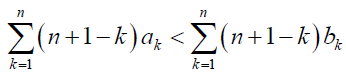 (6)
(6)
It is also interesting to investigate whether A is more likely to replace B than vice versa. In the limit of large N, by using (1)- (4), it turns out that ρA is large than ρB if and only if
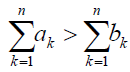 (7)
(7)
In the RESULTS section, we assume that the population size is sufficiently large, and we use (5)-(7).
Case 1:
In Case 1, we consider the case where a1−b1 is positive and ak−bk is constant for any k satisfying k>1 and negative. As defined in Introduction section, p* is the frequency of A at the unstable equilibrium in the conventional deterministic model.
By using (5), in our stochastic model, ρA > 1/N is equivalent to
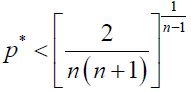 (8)
(8)
Note that this inequality gives the {2/[n(n + 1)]}1/(n – 1) law (a generalized version of the one-third law), obtained in Kurokawa and Ihara [4].
Similarly, by using (6), in our stochastic model, ρB > 1/N is equivalent to
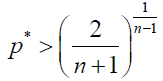 (9)
(9)
Similarly, by using (7), ρA > ρB is equivalent to
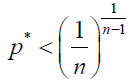 (10)
(10)
Note that these inequalities give the (1/n) 1/(n – 1) law (a generalized version of the one-half law), obtained in Ref [4].
Case 2:
In Case 2, we consider the case where a1−b1 is positive, and an−bn is negative, and ak−bk=0 holds true for any k satisfying 1 < k<n. Note that this situation occurs if additional fitness is given to every player when and only when every player adopts the same strategy.
By using (5), in our stochastic model, ρA > 1/N is equivalent to
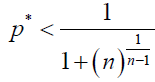 (11)
(11)
Note that this inequality gives another generalized version of the one-third law, obtained in Ref [3].
Similarly, by using (6), in our stochastic model, ρB > 1/N is equivalent to
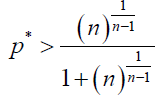 (12)
(12)
Similarly, by using (7), ρA > ρB is equivalent to
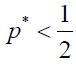
Note that these inequalities give the one-half law (a generalized version of the one-half law).
Stochastic dynamics as well as deterministic dynamics (such as replicator dynamics) is interesting. Nowak et al. [3] obtained the simple rule, called the one-third law. Kurokawa and Ihara [4] extended the one-third law and obtained the {2/[n(n + 1)]}1/(n – 1) law; however, when this law holds true is not explored. This paper examined when the law ({2/[n(n + 1)]}1/(n – 1) law) holds true and furthermore, we derive another extension of one-third law (1/[1 + (n) 1/(n – 1)] law).
Lessard [13] has revealed that (5)-(7) have been shown to hold also for an exchangeable model. Therefore, the findings in this paper are robust also for an exchangeable model.
This work was partially supported by Grant-in-Aid for JSPS Fellows and by Chinese Academy of Sciences President’s International Fellowship Initiative. Grant No. 2016PB018.