Research & Reviews: Journal of Botanical Sciences
ISSN: 2320-0189
ISSN: 2320-0189
Bel Hadj Chedli Rim1*, Abid Ghassen2, Sassi Khaled1
1Department of Agronomy and Plant Biotechnology, University of Carthage, National Agronomic Institute of Tunisia, Tunis-Mahrajene, Tunisia;
2Department of Sustainable Agrosystem, Centre University of Biotechnology of Borj Cedria, Hammam-Lif, Tunisia
Received: 02-Mar-2022, Manuscript No. JBS- 50710; Editor assigned: 04-Mar-2022, PreQC No. JBS- 50710 (PQ); Reviewed: 18-Mar-2022, QC No. JBS-50710; Revised: 25-Mar-2022, Manuscript No. JBS-50710 (R); Published: 01-Apr-2022; DOI:10.4172/ 2320-0189.11.3.001
Visit for more related articles at Research & Reviews: Journal of Botanical Sciences
The effects of the artificial inoculation by two biofertilizer based on Arbuscular Mycorrhizal Fungi (AMF) in the presence and absence of compost were studied on some morphological, physiological and agronomical traits of cultivated durum wheat (Triticum turgidum L. var. durum Desf.) in Northern Tunisia according to organic cropping system. Their effects were also investigated on the expression profiles genes involved in nitrogen assimilation: (GS) and (NR) encoding for Glutamine Synthetase and Nitrate Reductase respectively. Results indicated that the best grain yield, 1000 grain weight, grain number per spike, spike number per square meter and spike length, are obtained in plants that received the combined treatment (compost before planting and inoculations with AMF). This treatment significantly increased biological yield (9.1 quintals/ha against 7.2 quintals/ha in the control), chlorophyll (29.18 mg/g FM against 13.24 mg/g FM in the control), dry weight (7.1 T/ha against 5.1 t / ha in the control), plant height (76.61 cm against 66.6 cm in the control) and the leaf area (18.93 cm2 against 12.4 cm2 in the control). On the other hand, results indicated that an over-expression of (GS) was observed after the application of AMF and also in the combined treatment (AMF-compost) especially during grain filling stage. Higher expression of NR was also recorded in mycorrhizal-composted wheat plant after and before grain filling stages. Thus, this approach helped to reveal the importance of the application of mycorrhizae-compost in terms of improving plant growth and wheat production.
Wheat; Mycorrhizal fungi; Nitrogen assimilation; Biofertilizer
Agrochemicals are widely used in agricultural production to control pests, diseases, weeds, to improve yield and to maintain high product quality [1]. Despite their popularity and extensive use, agrochemicals constitute a real threat for humanity arising from the exposure of farmers when mixing products or working in treated fields, and also from residues on food and water [2]. The search for alternative and eco-friendly solutions is being an urgent need to find sustainable agriculture [1-3]. However, the application of molecular technique appears as useful tool for new research in sustainable food production and bio-product as well as to define the influence of micro-organisms to improve the cropping systems performances [4,5]. The introduction of plant growth-promoting microorganisms in crop production is considered as a good sustainable strategy to ensure competitive yields in many crops and to improve the resources use efficiency [6,7].
Among the beneficial microorganisms, the Arbuscular Mycorrhizal Fungi (AMF) belonging to the phylum Glomeromycota [8], are considered as the most beneficial fungus which could establish a symbiotic association with the roots of many plant species [9], including durum wheat (Triticum turgidum L. var. durum Desf.) [10]. These fungi not only promoted root growth and facilitated plant performance in drought environments [11,12] but also it was found that they could protect plants from abiotic and biotic stresses [13-15], improve soil quality and structure [16], increase water availability [12], enhance nutrient uptake such as Phosphorus (P), Nitrogen (N,) and Zinc (Zn) and increase grain yield by 20% [5,17]. Hence, it has been reported that wheat plants inoculated with AMF achieved higher growth, yield and nutrient uptake than non-mycorrhizal controls when grown in semi-arid areas [13]. Further, results of research studies in the field and in greenhouse using AM fungal inoculation of wheat showed a decrease of agrochemicals application, especially the application of Phosphorus fertilizer [1, 18].
On the other hand, it has been suggested that increasing organic matter in the soil will enhance the production of AMF biomass [1], and that organic amendments with high concentrations of Nitrogen (N) allows to increase AMF growth [19]. Recently, Hodge and store [20] reported an increased mineralization in the presence of AMF hyphae, which may suggest the availability of mineral nitrogen. Although the role of AM fungi in improving the phosphate nutrition of their host plants has been extensively studied [21,22], their contribution to the nitrogen assimilation is still controversially discussed [23,24]. The nitrogen is taken up from the soil by the AM extraradical mycelium [25]. NO3− is taken up by plants or fungi and reduced to nitrite by a Nitrate Reductase (NR) enzyme. Therefore, NR activity is mainly driven by NADPH in AM fungi and its activity in both roots and shoots of AM plants have been well documented [26,27]. The nitrogen form NH4+ is rapidly assimilated into amino acids according two pathways: the NAD(P)-Glutamate Dehydrogenase (GDH) or the Glutamine Synthetase-Glutamate Synthase (GS-GOGAT) pathway [20,24]. The GS and GOGAT activities have been reported to be significantly higher in roots and shoots of AM plants [26,27].
Keeping in view the significant role of mycorrhiza and compost in improving soil properties and crop production [28], up to now, not much is known about their combined role in nitrogen assimilation enzymes such as (NR) encoding for (Nitrate Reductase), and GS (encoding for Glutamine Synthetase) in wheat (Triticum turgidum L. var. durum Desf.). Thus, the aims of the present study were: i/-to evaluate the effects of beneficial fungi inoculations on, chlorophyll content, leaf area, plant length, biological yield and yield components of the Tunisian durum wheat ‘Maali’ cultivar, ii/ to study the role of combined AM fungus and compost before sowing in improving wheat production, iii/. To investigate the impact of AM fungi in synergy with compost application on the expression level of the plant nitrogen assimilation enzymes: NR and GS encoding for nitrate reductase and glutamine synthetase respectively.
Site description and experimental design
Field experiments were conducted during 2013–2014 wheat growing season at El Kawekib’ experimental station (36° 31′ N, 9° 58′ E, governorate of Zaghouan, northwest of Tunisia), located in the upper semi-arid bioclimatic zone where the annual rainfall reached 211 mm and the daily mean temperature varied between 4 and 34°C (during 2013-2014). The Tunisian durum wheat “Maali” cultivar was sown in the second half of December with a sowing rate of 400 viable seeds/m2 after a fallowed field. The experiment was conducted under rainfall conditions with complementary irrigation and no fungicides/pesticides were used to provide the crop protection. The Randomized Complete Block Design (RCBD) with three replications was adopted and a control treatment, without any fertilization, was then carried out. The plots were divided into 18 experimental plots with a surface area of 60 m2for each plot. The soil was clay with: pH:7, NH4+: 6.56 mg/kg, NO3-: 32.29 mg/kg, P2O5:16.11 mg/kg, K2O:21.05 mg/kg.
Arbuscular mycorrhizal fungal inoculum
The effect of two commercial biofertilizer based on AMF have been tested and six treatments were carried out under open field conditions and 18 sub-plots treatments were consisted of AMF inoculation. Two foliar and seed coating AMF biofertilizer were then used in this study (AMF 1 and AMF 2, respectively) in the presence and absence of compost application. Details of all treatments, their time of application and doses were represented in Table 1 . The composition of the commercial biofertilizer AMF1 was: (Glomus coronatum, Glomus caledonium) and for AMF2 was: (Glomus intraradices, Glomus mosseæ, Glomus viscosum).
Estimation of chlorophyll contents
Flag leaf samples were collected during the heading stage (GS58) for the determination of chlorophylls a and b contents using spectrophotometer according to Arnon (1949) calculations and cited by Cheikh M’Hamed et al. [29]
CH t=CH a+CH b
CH a (mg/l)=12,7 OD (663)-2,59 OD(645).
CH b (mg/l)=22,9 OD(645)–4,68 OD(663).
CH a: Chlorophyll a concentration
CH b: Chlorophyll b concentration
CH t: total chlorophyll concentration.
OD: Optical Density.
Flag leaf area, plant growth and yield components
Wheat flag leaves were sampled during the heading stage (GS 50 to GS 58). Leaf area was measured using a planimeter (LASER AREA METER CI-202). At the maturity stages (from GS 75 to GS 100), plant length, spike length, numbers of spike per square meter, grain number per spike, were recorded on the 10-plant samples from each plot. However, the above-ground biomass was oven-dried at 65°C for 48 h to determine the aboveground biomass during stage (GS 45). After harvest, 1000-grain weight, grain yield and the biological yield in each plot were also determined.
| Treatments | Composition | Time of application |
|---|---|---|
| T 1 | AMF 1 (20Kg/Ha) | Tillering and heading stage |
| T 2 | AMF 2 (20Kg/Ha) | Seed coating |
| T 3 | AMF1+AMF2 | Seed coating with AMF2, and AMF1at tillering and heading stage |
| T 4 | Compost (9T/Ha)+AMF2 | Compost before sowing and seed coating with AMF2 |
| T 5 | Compost +AMF1+AMF2 | Compost before sowing, seed coating with AMF2 and AMF1 application at tillering and heading stage |
| T 6 | Uninoculated control | - |
Table 1. Treatment combinations based on AM tested during the experiment.
Statistical analysis
Analysis Of Variance (ANOVA) was performed for each treatment using the SAS, [30] Least Significant Differences (LSD) at P ≤ 0.05 were considered significant. Mean values are presented in the Table 2.
| Number Accession in GenBank | gene | Length (bp) | Identity | % Identity (bp/bp) | E-value |
|---|---|---|---|---|---|
| BF293807 | GSP | 450 | Triticum aestivum alpha-gliadin storage protein U51306.1 | 438/450 (97%) | e-4 |
| AJ611392 | NR | 330 | H.vulgare mRNA for nitrate reductase X57844.1 | 243/317 (77%) | 5.00E-57 |
| AJ609930 | GS 1 | 351 | Triticum aestivum glutamine synthetase isoform GS1 (GS) gene AY491968 | 317/336 (94%) | 4.00E-147 |
Table 2. Identification of GS, NR and GSP on durum wheat « Triticum turgidum L. var. durum Desf. »
Nitrogen genes assimilation expression levels
Plant materiel: Flag leaves were collected during three growing stages (according to Zadok stages 1974): 1/- Booting stage (GS 47: Flag leaf sheath opening) 2/- Grain filling stages (GS70) and 3/- at the end of ripening stages (GS99). However, for each stage, samples were ground immediately in liquid nitrogen, and stored in -80°C.
RNAs extraction: Total RNA from wheat leaves, was extracted according to Chang et al. [32] method with quite modifications. Thus, approximately 200 mg are ground in liquid nitrogen and then transferred into a sterile eppendorf tube to which 750 µL of extraction buffer (2% CTAB, 2% PVP, Tris-HCl 100 mM pH 8, 25 mM EDTA, 2M NaCl, 0,5 g/l spermidine, 15 µl of ß-mercaptoethanol) were added. After a vortex mixing, 750 µl of chloroform/isoamylalcohol (24/1) were also added. After 10 min of centrifugation at 13000 rpm, the supernatant obtained was then transferred into a sterile eppendorf to which 200 µl of LiCl was added and the tubes were kept at 4°C overnight. After 30 min of centrifugation at 10000 rpm, the pellet ARNs were added to 200 µl of DEPC treated water. To precipitate RNAs, 50 µl of NaAc 3 M pH 5, 2 and 500 µl of ethanol were added and the tubes were kept at -80°C for 1 h. After 30 to 45 min of centrifugation at 10000 rpm, the pellet RNAs were washed too times with 70% cold ethanol. The pellet was dried in a flow-cabinet and then suspended in 100 µl of DEPC treated water. The RNA was quantified at 280 nm ((UV-VIS Dual Beam).
RT-PCR: RT–PCR was carried out using ‘RevertAid First Strand cDNA Synthesis (Fermentas) kit’.However, 10 µg of total RNA was treated with 10 Uof DNaseI (Fermentas) in DNAseI assay buffer for 30 min at 37ºC. Four microgram of total RNA was reversed transcribed into cDNA by random priming with the RevertAidTM H Minus M-MuLV RT 200 U/μl (Fermentas) at 42ºC for 1 h according to the manufacturer’s instruction. Fragments were amplified from these cDNA pools using Taq DNA polymerase (Biolab’s), following the manufacturer’s procedure.
The 18S gene was used as a control for RNA loading. PCR amplification was performed for all genes using the following program: 3 min at 94C, 30 s at 94C, 45 s at 55-60C and 1 min at 72ºC. An elongation step at 72ºC for 5 min was conducted for the last cycle. Reactions were performed in GeneAmp® PCR System 9700 (Applied Biosystems) thermal cycler.
Database searches: In silico analysis identified specific nucleotide sequences corresponding to the genes of interest (GS and NR) and reference gene (18 S) in durum wheat in the National Center gene library for Biothechnology Information "(NCBI, http://www.ncbi.nlm.nih.gov) as shown in Table 2. For this research, the Basic Local Alignment Search Tool, PSI-BLAST program (http://www.ncbi.nih.gov/blast/) was used to identify the sequence in durum wheat with similarity to the input sequences in other species (Arabidopsis thaliana, Triticum aestivum…).
Polymerase chain reaction analysis
The amplification was carried out using ‘Biomatik kit’. As a control, the housekeeping gene Pv 18S. Table 3 was amplified. PCR was conducted in 20 μl of reaction volume containing 0.5μl of each primer, 1 μl of cDNA template, 2 μl PCR buffer (10x), 2 μl mix dNTP, 2 μl MgCl2 and 0.2 μl Taq DNA Polymerase following the manufacturer’s procedure, under the following conditions: 94°C for 3 min, and 40 cycles of 94°C for 30 s, 59°C for 45 s, and 70°C for 1 min. Real-time PCR reactions were carried out in GeneAmp® PCR System 9700 (Applied Biosystems) thermal cycler. The amplified PCR products (25 μl) were electrophoresed on 1% agarose gel, stained with ethidium bromide, and then scanned using an image analyzer UV (Transilluminator).
| Gene | Forward primer 5’-3’ | Reverse primer 5’-3’ | Tm |
|---|---|---|---|
| 18S | F-TCGCATCGATGAAGAACG | R-CTTGAGAGACGAGGGACC | 58°C |
| GSP | F-CCACAGTATTCGCAACCA | R-TTCCATGCGCTATGTTGT | 61°C |
| NR | F-CGATGACACCACGGTAGATG | R-CACCACGTACCACACCTTGA | 61°C |
| GS 1 | F-GAAGGTTGCAGCTGAGGA | R-CGTAGAGGCAGGCCTTGTAG | 60°C |
Table 3. Primer sequences used for quantitative real-time PCR.
Effect of mycorrhizal inoculation on morphological parameters of durum wheat
Plant and spike length: Results of this study showed that the bio-inoculant T1, Table 1 have a significant effect (P<0.05, Table 4) on plant length, while T3 and T4 Table 1 did not produce positive effects on this morphological parameter as shown in Table 4. Wheat plants from the coated seeds T2, Table 1 displayed a significant increase (P<0.05, Table 4) in plant length, compared to those from the uncoated seeds treatment. However, results also clearly indicated that the plant length was significantly 1%, Table 4 influenced by application as well as combined treatment T5 (Compost+AM1+AM2, Table 1 ) with the highest mean plant length (76,61 cm) followed by 70.95 cm and 70.71cm for treatments T2 and T1 respectively (Figure 1).
| Treatments | ||||||
|---|---|---|---|---|---|---|
| Parameters | Absence of compost | Presence of compost | ||||
| T1 | T2 | T3 | T4 | T5 | ||
| Plant length (cm) | F value | 27.99 | 2.27 | 3.57 | 7.05 | 16.29 |
| Pr>F | 0.0018 | 0.0614 | 0.1077 | 0.1267 | 0.001 | |
| spike length (cm) | F value | 4.12 | 4.38 | 0.34 | 8.28 | 9.76 |
| Pr>F | 0.665 | 0.0812 | 0.582 | 0.034 | 0.0291 | |
| Leaf area (cm) | F value | 8.52 | 9.81 | 1.12 | 1.14 | 26.48 |
| Pr>F | 0.0429 | 0.0273 | 0.3298 | 0.3858 | 0.001 | |
| Chlorophyll contents (mg/g FM*) | F value | 11.48 | 3.81 | 47.72 | 1.7 | 13.98 |
| Pr>F | 0.0147 | 0.253 | 0.0005 | 0.2596 | 0.0002 | |
| Biological yield (T/Ha) | F value | 1.1 | 8.71 | 0.19 | 9.98 | 3.74 |
| Pr>F | 0.3347 | 0.0256 | 0.8033 | 0.013 | 0.35 | |
| Biomass maturity stage (T/Ha) | F value | 1.25 | 4.94 | 1.92 | 21.9 | 0.75 |
| Pr>F | 0.3064 | 0.0679 | 0.2264 | 0.0034 | 0.4776 | |
| Spike number /m2 | F value | 2.71 | 2.8 | 6.33 | 9.55 | 7.98 |
| Pr>F | 0.12 | 0.1214 | 0.047 | 0.021 | 0.04 | |
| Grain number/Ear | F value | 1.78 | 3.27 | 1.15 | 0.02 | 1.25 |
| Pr>F | 0.23 | 0.1204 | 0.3244 | 0.9761 | 0.3906 | |
| 1000 grain weight (g) | F value | 2.81 | 4.59 | 0.49 | 0.46 | 12.66 |
| Pr>F | 0.1446 | 0.179 | 0.5119 | 0.6537 | 0.015 | |
| Grain Yield (quintals/Ha) | F value | 5.82 | 13.12 | 70 | 85.95 | 110.62 |
| Pr>F | 0.0525 | 0.0123 | 0.0002 | 0.0001 | 0.0001 | |
Table 4. ANOVA values of measured morphological, physiological and agronomic parameters.

Figure 1: Effect of individual and combined treatments on plant length. 
A significant effect (P<0.05, Table 4) has been observed after the application of the combined treatment T5 and T4 Table 1 with the highest mean spike 15.01 and 14.74 cm respectively against 13.2 cm in the control (Figure 2).

Figure 2: Effect of individual and combined treatments on spike length. 

Effect of mycorrhizal inoculation on physiological parameters of durum wheat
Flag leaf area: The combined treatment T5 improved significantly (1%, Table 4) the flag leaf area which reached the maximum (18.81 cm2, Figure 3) compared to the control (12.61 cm2, Figure 3). There were also a clear differences in the application of the two biofertilizer AM1 and AM2 in the absence of compost (T1 and T2 respectively, Table 1 ).

Figure 3: Effect of individual and combined treatments on leaf area. 
Chlorophyll contents: The total leaf chlorophyll contents (chlorophyll a and b) were significant in the treatments T3 and T5 (P<0.001, Table 4) with 26.09 and 25.49 mg/g FM in T3 and T5 respectively (Figure 4). The treatment T1 increased also the chlorophyll content by 21.63 mg/g FM compared to 14. 43 mg/g FM in the untreated control.

Figure 4: Effect of individual and combined treatments on chlorophyll contents. 

Crop biomass and biological yield: The biomass at tillering stage and the biological yield were insignificant in (T1, T3, and T5). In contrast, a significant effect was noted after T2 and T4 application where more than 7.55 T/ha and 8.76 T/ha (Figures 5 and 6) of biomass at tillering stage and biological yield were recorded respectively against 6.96 T/ha in the control.

Figure 5: Effect of individual and combined treatments on biomass at tillering stage. 
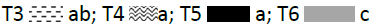

Figure 6: Effect of individual and combined treatments on biological yield. 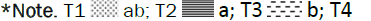
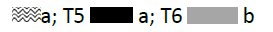
Effect of mychorrizal inoculation on yield components
Analysis of variance ( Table 4), indicated that the number of spike/m2 increased significantly (P<0.05) in the treatments T4, T3 and T5 with 294, 300 and 303.67 spike/m2 respectively (Figure 7). Thus, no significant difference was noted for grain number/spike neither in the presence nor in the absence of the compost. Hence a very quiet increase of this parameter from 29.16 grain/spike in the control to 34.22 grain/ spike in the combined treatment T5 Figure 8 have been recorded. Same trend has been observed with 1000-grain weight where only T5 treatment showed a significant increase (P<0.05, Table 4) for this parameter (from 50.95g in T5 against 43.98g in the control, Figure 9). These results indicated also a significant increase 1%, Table 4 of grain yield in T5, T4 and T3 treatments which reached 30, 27, 25 quintals/ha respectively against 16.04 in the control untreated (Figure 10).

Figure 7: Effect of individual and combined treatments on spike number/m2. 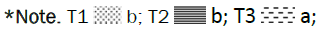


Figure 8: Effect of individual and combined treatments on grain number/spike. 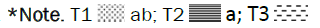


Figure 9: Effect of individual and combined treatments on 1000-grain weight. 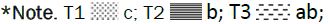


Figure 10: Effect of individual and combined treatments on grain yield. 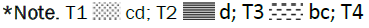
Effect of am fungi inoculation on nitrate reductase (nr) gene expression
Results of this study indicated that the NR expression level was higher during booting stage (GS 47: Flag leaf sheath opening) compared to the grain filing and ripening stages as showed in Figure 11. Thus, before grain filling (booting stage) the Nitrate Reductase gene (NR) was over-expressed after the individual inoculation of the two biofertilizer T1 and T2, Table 1 and also in the combined treatment T4 compared to the control untreated T6 (Figure 11). Moreover, in ripening stage, only the treatments T1 and T3 seems to improve the NR expression level as presented in Figure 11.

Figure 11: Gene expression profiles for Nitrate Reductase (NR) at booting stage GS 47 (A) Grain filling stage GS70 (B) and ripening stage GS99 (C).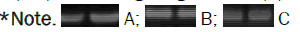
Effect of am fungi inoculation on glutamine synthethase (gs) gene expression
The real time PCR that verified the differences in gene-expression between treated and untreated plants, showed a positive effect of the use of the two fertilizer T1 and the combined treatment T4 ( Table 1 ) in induction of expression nitrogen genes assimilation GS during seed development stage (Figure 12) compared to the control untreated. The GS profile expression still higher also during the ripening stage in T2 and in the combined treatments T3 and T5 ( Table 1 ) while no effect of AFM inoculum has been noted at booting stage (Figure 12). This increase in gene expression may be credited to mycorrhizal inoculation as it resulted in improvement of Nitrogen uptake. Results obtained in this research showed a weak expression of Glutamine Synthase (GS) gene before the filling phase while a clear overexpression was observed during and after seed development (Figure 12).

Figure 12: Gene expression profiles for Glutamine Synthetase (GS) at booting stage GS 47 (A) Grain filling stage GS70 (B) and ripening stage GS99 (C). 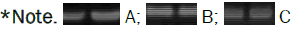
The exploitation of Arbuscular Mycorrhizal (AM) fungi has become of great interest in agriculture due to their potential roles in reducing the need for agrochemicals and improving plant growth and nutrition in a sustainable and environmentally way [1]. In this study we investigated the effect of the combined treatment AMF bio-inoculants and compost on the organic durum wheat production under field condition.
Results showed a significant difference of these treatments for the majority of the measured parameters. The most important plant length mean was observed in the combined treatment (T5) and also after the application of the two biofertilizer separately which could be mainly associated with to the great increase of nutrient uptake regulated by the AMF [33,34]. These results are in agreement with previous studies which reported that mycorrhizal inoculations enhance plant length in barely and in durum wheat by coating seeds fungi [17, 35]. Moreover, the two tested biofertilizer when applicated separately, increased the flag leaf area in presence and absence of the compost and a significant performance combination of the mycorrhizae and compost (T5) was obse37rved on leaf area. These results indicated that the compost application affects mycorrhizae in plant nutrition, renders higher nutrients uptake as well as improved soil fertility [36, 37].
The application of the seed coating AM2 inoculation without compost (T2) enhanced the chlorophyll contents, the biomass during tillering stage and the biological yield in durum wheat plants. Similar results were revealed by Hussain et a1l. [28], Moucheshi et al. [38] and Oliveira et al. [1], confirming the increase of chlorophyll content and the photosynthetic activity in inoculated plants of barley, corn, soybean, lettuce under conditions of phosphorus deficiency [39]. Great increase of biomass and biological yield after AMF inoculation on barely, maize and durum wheat respectively were then observed in many previous studies [40-42]. The combined treatment AMF and composts seems to have a great effect on the chlorophyll contents, the biomass during tillering stage and the biological yield in this experiment as a result of nutrient uptake enhancement after the compost application which has been shown to be highly efficient in association with arbuscular mycorrhiza [11].
In this study, inoculation not only enhanced the physiological traits of durum wheat, but also influenced the yield components. Thus, the improved performance after AMF inoculation for spike length, spike/m2, grain number/spike, 1000-grain weight and grain yield in field conditions was probably due to the absorption of more nutrients by wheat plants and the enlarge of the effective surface outside of the roots [18,36-39]. In this context, [43] showed that AMF symbiosis in maize enhances spike dry weight, spike length, spike circumference, the dry weight and dimensions of the grain. Furthermore, [5] and [1] reported that AMF inoculation increased biomass and grain yield by 20%, showing that productivity could be increased by applying inoculum in the field.
However, the highest yield components (grain yield, 1000 grain weight, grain number per spike, spike number per square meter and spike length), are recorded for dur5um wheat received the combined treatment (compost before planting and inoculations with AMF). This increase may be related to the mycorrhizal inoculation in synergy with compost application before sowing as it resulted in improvement of phosphate uptake [11-20,44]. Several studies have demonstrated that crop performances increased with AM fungi in synergy with organic amendment [11] and vermi-compost [28]. In contrast, [45] and [33] suggested that the grain yield of durum wheat was not affected by AMF inoculation under fertilization.
Overall results recorded suggest that AMF-compost association plays an important role in morphological, physiological and agronomic traits. The improvement of the plant N nutritional status may be the origin of the increase in growth of durum wheat plant. The obtained data indicated a high level of NR gene expression was revealed on durum wheat inoculated flag leaves after the use of two biofertilizer based on AMF as a result of the increases in activities of key N-assimilating enzymes (NR). In a recent review, [24] reported that NR activity is mainly driven by NADPH in AM fungi and its activity in shoots of AM plants is generally higher than in non-mycorrhizal control plants. Similar results confirmed the improvement of NR activity in AM-shoots of maize plant (Zea mays L.) compared to non-AM shoots [26, 46].
Furthermore, a high level of NR expression gene after the use of the combined treatment T4 (compost+AM2) was noted especially before grain filing and in ripening stages. The synergy application of compost-AM fungi which enhance plant nutrient could explain the increase in NR activity in durum wheat flag leaves. Previous studies conducted by [47, 48], revealed a synergistic effect between the fermented amendment and AM fungi in increasing NR activity in roots of J. Oxycedrus and D. pentaphylum. Hence,the combining effects from the NR activation through nitrate addition via organic amendment with improved P uptake through mycorrhizal fungi enhance the N assimilation of the plant [11].
The ammonium originating from nitrate reduction is mainly assimilated in the plastid/chloroplast by the so-called GS/GOGAT cycle and the Glutamine Synthetase (GS) fixes ammonium on a glutamate molecule to form glutamine [49, 50]. Results obtained in this research showed a weak expression of Glutamine synthtase (GS) gene before the filling phase while a clear over-expression was observed during and after seed development (Figure 2). Therefore, a strong expression of this gene has been observed in previous research during grain filling phase [51] and senescence [52] in maize (Zea mais).
However, the most important GS gene expression level was observed in treatment based on AMF (T1) during filling stage which could may be credited to mycorrhizal inoculation as it resulted in improvement of Nitrogen uptake [13]. The GS gene expression in wheat leaves is consistent with those found in maize shoots (Zea mays L.) when colonized with Glomus fasciculatumn [26]. Early study conducted by Subramanian and Charest [46] reported that GS as well as GOGAT activities were higher on mycorrhizal maize shoots compared to the untreated ones. In another context, mycorrhizal treatment increased GS activity in mycorrhizal plants compared to control plants of Dendrobium chrysanthum after 90 days of inoculation [53]. Great expression level of GS was noted also in almost combined treatments (AFM and compost) during filling grain phase. The same trend was observed in ripening stage in T3, T2 and T5 treatments. The higher nutrient supply arising from the compost and as well as the AFM nutrient uptake such as phosphate and nitrogen uptake could explain the increase expression gene level of GS recorded in these experiments [20, 44].
Overall, the higher expression levels relative to GS and NR in both, mycorrhizal wheat plant, might be due to assimilation of NH4+ ions derived from the primary NO3- reduction as well as other metabolic pathways, including direct uptake, photorespiration and amino acid catabolism [44]. However, it has been demonstrated in the early studies, that the Glutamine Synthetase (GS) is the main responsible enzyme for N assimilation by AM fungi [54]. Besides, the Nitrate Reductase enzyme (NR) has been identified in the nitrogen assimilation pathway so far in44 these fungi. Nevertheless, few authors have highlighted the importance of mycorrhizal association on the GS activity in organic durum wheat. Yet, despite the proven benefits of AM fungi on wheat, there is no study available for their application in combination with organic amendment on GS as well as NR under open field condition [55].
The work presented here is a first step in filling the gap in the knowledge of the effect of the combination AM fungi-compost on N assimilation gene expression before, during and after filling stage. A genetic characterization has validated the beneficial effect of the combined mycorrhizal inoculation and compost application on nitrogen gene assimilation in organic durum wheat before, during and after grain filling. A positive effect of the application of the combined treatment AMF-compost in terms of improving plant growth and wheat production was further demonstrated. Future research should focus on the importance of this specific response many critical questions are still unanswered and should be addressed in future studies.
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]
[Crossref] [Google Scholar] [Pubmed]