Research & Reviews: Journal of Ecology and Environmental Sciences
ISSN: 2347-7830
ISSN: 2347-7830
Institute of Biochemistry and Biotechnology, University of the Punjab, Lahore, Pakistan
Received Date: 31/08/2017; Accepted Date: 27/09/2017; Published Date: 06/10/2017
Visit for more related articles at Research & Reviews: Journal of Ecology and Environmental Sciences
Drought is an environmental restraint for plants which would become more severe in coming years if effective measures won’t be taken. It is a condition of reduced or very low availability of water to plants for a prolonged period of time hence disturbing plant growth, development, water relations, and efficiency of many terrestrial plants. Plants acclimatize themselves to different sophisticated biochemical, physiological, morphological changes to overcome drought conditions. Crops are prone to high susceptibility towards decrease in grain yield, reduced transpiration, and changed hormonal balance regulated through mechanisms of activation of anti-oxidant defense system, stomatal pore regulation, and osmoregulation in plant tissues. With advances in drought tolerance and plant genetic engineering, many drought tolerant traits can be acquired
Drought tolerance, Leaf area index, Water use efficiency, Physiology, Plant regulation
Plants are affected by drought conditions which is one of the major problems faced by the plants. Methods are being employed to increase the Water use efficiency (WUE) for plants. By altering the irrigation time and genotyping of plants we can manage the amount of water used by the crops [1]. Plants usually begin various mechanisms to sustain water in them such as through formation of deep roots, pumping of salts into the vacuole and out of the cells, increase of water resistive diffusion, and the formation of small succulent leaves to reduce water loss [2]. Osmolytes including polyols, metabolites, organic acids, glycinebetaine, and proline along with many other aminoacids play an important role during drought conditions to sustain the proper functioning of plant cells [3]. There are many enzymes and substances like polyamines and citrulline which work as effective antioxidants to limit the effect of drought [4]. Researchers have developed several techniques to make plants resistant towards drought conditions through breeding and massive screening, application of phytohormones exogenously to the plants, subjection of some osmo-protectants to the seeds for germination and good growth, genetic engineering, and marker based selection methods [5]. The environmental conditions leading to drought include low precipitation and high rate of evapo-transpiration causing reduction in plant cell division, proliferation of roots, closure of stomatal pores, changes in relation between plant and water uptake efficiency, and high production of Abscisic acid which further helps decreasing the evapotranspiration by controlling stomatal pores [6]. Since plant growth is a reversible process with myriad metabolic pathways, all the phases such as development, growth, differentiation of cells, division, elongation, increase in volume, and intensification of weight are irreversible in it and get affected [7] . In this review paper we have highlighted the advances which are being made to make plants more droughts tolerant; starting from traditional methods gene and regulation, genetic engineering, and plant physiology along with numerous essential substances which play an important role in plant metabolism.
Traditional Breeding Methods for Drought Tolerance
As for the need to make plants more drought tolerant, we need to get the specific variety of the plant which shows best results for the plants which are best tolerant, thus the varieties at the inter-specific, intra-specific and inter-generic levels are taken into consideration [8]. Dr. Norman Borlaug and their co- workers found the breeding lines which were found to be produced at the best for disease resistance and pest resistance, two decades were passed to get the breeding line. Among sexually compatible plants, these crosses were made to find the result. Wheat, the main staple food was found to be badly affected by the drought. Similarly, the international rice research institute was found to improve the quality and yield of rice. Conventional breeding rather becomes impossible if the proper genetic variation in the gene pool becomes considerably low. Marker assisted breeding have been shown to allow us the study of the plants with different genetic combinations. It allows the breeder to assist gene pools to control specific phenotypes. Genetic engineering has been in practice to produce transgenic crops which show high tolerance to the biotic and abiotic factors [9]. As an example, rice plant containing the chimerical gene Ubi1. TPSP was found to show the increase in the drought tolerance due the increased production of the trehalose [10].
Recent Advancements in Drought Tolerance
Scientists have compared the crops by measuring the working capability through water requirement expression. The linear relationship was found for the water requirement and dry mass produced. It was found that water efficiency was not affected by the soil water. Severe climate, difference of plant species, and the availability of soil mineral were among the major reasons. It has been observed that differences between C3 and C4 species were effectively prominent. Tests were not done in species exhibiting Crassulacean Acid Metabolism. Considering the example of pineapple having similar metabolism where it concentrates CO2 to organic acids at night and causes it to release the next day for the process of photosynthesis. In this mechanism, the stomatas are found closed and water is conserved. This allows CAM plants to achieve higher water saving than C4 plants [11]. Mathematicians and Scientists have now explained the concept of water use efficiency or WUE which is the total amount of the dry mass produced by the plants by using per unit of water.
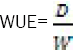 (1)
(1)
Where, D is the mass of the plant considered to be dry while W is the amount or mass of the water used.
Alfalfa plant utilizes less water than maize when conditions are not favorable. Economically, there would be preferable options with rise in the level of water to change the force. Dry mass of the plants majorly involve carbon and oxygen. These are heavier than hydrogen atoms, which originates in the process of hydrolysis of water in the photosynthetic processes. The waxy cuticle and stomatal opening prevents loss of water, through controlled opening and closing of stomata in specific conditions. Water use is also affected by environmental factors like humidity, temperature, air, and CO2 concentration. For the reduced amounts of humidity, transpiration rate increases resultantly [12]. Plants with improved drought tolerance were produced, while it was a difficult approach as the plants need water as a basic need, but it was made possible [13]. Improved grain plant which shows the increase in drought tolerance by testing the varying location and different availability of the water by making crosses to get the most tolerant species to show high yield in the deficient conditions. But they need no sacrifice under favorable conditions. Experimental results show that there could be advantage of osmotic adjustment without sacrificing of the amount of yield under optimized conditions. Importance of deep rooting was also inferred by that the deep rooted trees show more tolerance than the grasses with small less penetrating roots. While cuticle play a vital role in the tolerance increasing of the plants the variation was studied for the plants when they have stomata on both sides relating it with the thickness of cuticle layer. The amount of wax in rice was less than the paddy rice. In USA epicuticle wax was deposited on the grasses to make it tolerant [14].
Molecular and Biological Regulatory Network
In response to drought conditions there is an alternation of gene expression and accumulation of osmotically active metabolites in plants. Phytohormone abscisic acid, oxygen scavenger radicals and specific chaperone proteins are being synthesized excessively [15]. Plant activates a diverse network consisting of metabolic, defense and physiological systems. Molecular metabolic system comprises a combination of regulatory and transcriptional factors, signaling proteins, and also functional genes working together. Microarray technology exposed that the TFs (transcriptional factors) such as B-zip, MYC, AP2 and ERF, MYB, and WRKY controlling the expression of downstream genes [16]. Transcriptional activators enhance the expression of drought tolerant genes during drought through their over expression. For example in Arabidopsis proteins which binds an element responding dehydration and CRT binding factor (DRBP/CRF3) acts as an activator. Transcriptional repressors favorably restrict or lessen the expression of some genes during stress conditions. In Arabidopsis AtMYB60 regulates stomatal movement by repressing the transcription of R2R3-MYB. Signaling factors signal protein phosphorylation and de phosphorylation, calcium sensing protein degradation, phospholipid metabolism and many other drought tolerance enhancing mechanisms. Through signal transduction system signaling factors are involved in various downstream processes [17]. In tobacco constitutive activation of a mitogen-activated protein kinase3 (MATKKK) NPK1 in response to oxidative signal cascade results in heat, cold, drought tolerance and salinity in transgenic plants. Signal transduction system is a complex system as there can be more than one output of signaling proteins. Combined functions of two signaling cascades of AtMPK6 (MKK2-MPK6 AND MKK4/MKK5 MPK6) transduce and transmits various signals from heat, cold and bacterial pathogens [18]. Functional genes control compatible solute synthesis and organic compounds synthesis i.e., amino acids, sugar, and sugar alcohols e.g. galactinol, mannitol, quaternary and other amines. ABA is a plant hormone being synthesized to sustain drought stress coded by a gene NCED3. In addition there is cytochrome family gene P45O CYP707A encodes ABA 8’-hydroxylase to control ABA levels in drought stress conditions and during seed imbibition. Overexpression of late-embryogenesis abundant (LEA) class of stress responsive proteins increases dehydration tolerance by preventing protein aggregation in being induced by freezing as well as drought. Its function is similar to chaperon which acts against cellular function [19]. Physiological system involves biochemical variations at the cellular level i.e., changing in fluidity of cell membrane, turgor pressure, and interaction of proteins with lipids and other proteins and solute concentration. Avoiding dehydration or tolerating dehydration controls turgor pressure. This in turn is regulated by morphology of roots (thickness, mass and depth of root) and developmental traits (compacted soil penetrating ability of root). Phenotypic traits are significant even at normal environmental conditions whereas dehydration tolerance and osmotic adaptation appears only in drought stress. Photosynthetic activity reduces and carbohydrate metabolism changes. Osmo-protectants give defense by protecting and maintain structure and function of membrane and cellular components. Reactive oxygen species are oxygen scavengers and anti-oxidants. Accumulation of Glycine betaine under stress conditions protects halophilic MOs. Quaternary ammonium, amino acids, tertiary sulfonium substances and polyols are some other osmoprotectants. Polyhydroxy compounds interacts hyrophobically through their hydroxyl group with membrane phospholipids. This interaction stabilizes membrane [20].
Engineering Drought Tolerance in Plants
Functional and regulatory genes directly involved in all stress related responses in drought tolerant plants are being engineered to enhance their function and to introduce these responses in drought non-tolerant plants. Arabidopsis is a model plant for studying drought tolerance because of availability of its complete genomic sequence, short term generation times, whole transformation protocols, proteomic data, microarray, EST sequence tags, and wide variety of mutants. Potential target gene for drought tolerance is selected in a rational way. Water, cold and salinity stress are treated as being correlated [21]. Microarray geneexpression technology is mostly used for identifying target gene. Genome-wise responses are analyzed by contribution of gene ontology. Due to complex nature of this drought stress response modification of single component or gene will not be enough. Several regulatory and signaling factors should be manipulated together. Cross-tolerant nature of target gene can be checked by additional abiotic stress response expression describing experiments. Microarray experiments also identify co-regulated genes. Up-stream promoter sequence is required for knowing cis-elements and transcriptional factors. Several databases serve this purpose. Transcriptional profiles vary between roots and shoot so crude overexpression of selected gene is not sufficient. Drought stress responses are function of tissues specificity and timing and vary widely. It will be beneficial to know about precise transcriptional changes occurring after drought to select and design target [22]. At last mechanism and site of expression of transgene is also necessary to know. Signaling molecule manipulation is a reproducible attempt but can result in unfavorable broad response. It is economical to manipulate stress-activation mediated signaling component instead of using overexpression of target gene. Engineered APETALAL2 transcription factor gene enables alfalfa (Medicago tranculata) to tolerate drought stress through wax production [23]. Members of R2R3-MYB gene of Arabidopsis AtMYB60 and AtMYB61 encode transcriptional factors controlling stomatal movement. Mutant Arabidopsis osm1/syp61 leads to impaired ABA-mediated stomatal opening. Engineered AtMYB60 gene through null mutation reduces stomatal opening [24]. In Arabidopsis post-transnationally activation of SRK2C kinase in response to stress enhances tolerance. In transgenic plants with engineered SRK2C gene this activity was observed much more. Through post-genomic tools transgenic constructs can be evaluated. Recently microRNAs responsive to drought stress have been discovered and used successfully. Mannitol dehydrogenase (MtlD) gene when introduced in wheat increased drought tolerant. In tobacco plant overexpression of transgenic inositol Methyl Transferase gene (IMTI) gene originated from ice plants enhanced drought tolerance [25].
Genetic Analysis and Physiological Interpretation
When drought conditions are met, plants acclimatize themselves by expressing surplus genes which shuts down the normal functioning and metabolism of cells. This happens when water reaches extremely low level raising an alarming situation for plants to produce a myriad of proteins of undefined functions. Many such proteins include de-hydrins as heat shock proteins and storage vegetative proteins [26]. There is a huge diversity of adaptation processes in plants. This is due to several environmental factors which effect plants such as salinity, deficit of water, isolations, heat shocks, and low-air hygrometry which creates different types of droughts. For variety of drought types, there are numerous mechanisms and metabolic reactions which take place at different levels of organizations. Of many physiological changes, the decrease in Leaf-area-index (LAI) due to suppression in leaf expansion contributes to reduced biomass and decreased seed growth yield [27] have described how plants such as rice, soya bean, and barley rapidly shift to the maturity phases and flowering on the onset of drought conditions. Conditions with early adoption of phase change often leads to the sterility of pollens of plants giving way to very poor plant crops yield. Another reflective change in the physiology includes the change in rate of influx of CO2 and nutritional uptake due to transpiration. Due to decrease in transpiration, the rate of photosynthesis decreases commendably along with the decrease in internal CO2 of leaf and reduced ATP production leading to reduced carboxylation. All the nutrients present in the plant cells require water for effective solubility and translocation of molecules. Downregulation of plant metabolic reactions reduces the substance Ribulose-1, 5-Bisphosphate or RUBP which also serves in carboxylation and photosynthetic utilization of CO2 for food. Rubisco also plays an important role in diminishing the high amount of internal CO2 and O2 which accumulates due to the closure of stomatal pores to increase the process of photo-respiration. It has been observed that there is severe decrease in Phosphorus, Nitrogen, and Potassium In shoots and roots of plants due to drought conditions [28]. There are many ongoing mechanisms in drought setting; including great production of Reactive Oxidation Species like radicals of Superoxide, Alkoxy Radicals, Single Oxygen, and Hydrogen Peroxide. These radicals cause high oxidative damage to the plant cells by destroying biomolecules of great importance such as proteins, and lipids. Destruction of Proteins and lipids further damages the protective functioning of plant cells. To nullify the effect of these radicals, many plants release superoxide dismutase, peroxide, catalase, and ascorbate peroxidase enzymes. Together, these enzymes prevent damage which occurs to the plants. There are many compatible solutes present in plants which never disturb cell membrane, plant enzymes, and biomolecules even if they are found to be in excess and such molecules include proline, sugar, organic acids, sugar-alcohol, and glycine-betaine or GB. These molecules protect the membranes and enzymes from the reactive oxygen species. The effect of glycine betaine is in protection of enzymes, membranes, and maintenance of the electrons flowing through the thylakoid membranes [29]. Many genotypes of the plants formulate mechanism to overcome drought through Drought Escape. To prevent the effect of extreme conditions of drought, they start producing seeds and spores during the period of short rainfall seasons to overcome extinction. Some plants have a genotype which leads to root plasticity and the increase of different favorable root features such as high proliferation of roots and increase in the depth and length of roots. Some plants have specific genotypes for osmotic adjustments, controlled phytohormonal dynamics, and up rise of defensive mechanisms against radicals [30].
Role of Aquaporins and Defensive Proteins
A wide range of mechanism and molecules have been studied which play important role in drought tolerance. Under the stress conditions, plants build up some new protective proteins like (HSP) heat-shock proteins and late embryogenesis abundant (LEA) proteins. Complete information regarding the regulation of these protective proteins can help biotechnologist to enhance the adaption and drought tolerance ability in plants. Under abiotic stress a group of proteins i.e., aquaporins are responsible for water regulation in the plants. Functional characterization of these aquaporins and other protective proteins has proved the importance of their regulation under abiotic stress. Food shortage is a worldwide problem occurs due to abiotic stresses and high population. To tackle this problem scientists go for gene revolution, which involves the transgenic approaches, following green revolution. For the agricultural yield, drought is one of the major restraints. To impart the crop resistance properties to the plants is a challenge for agricultural biotechnology. Currently the scientists are transferring one or more genes to achieve this standing goal. Overproduction of the protective genes in transgenic plants not only enhances the drought tolerance in plants but also improve the tolerance against abiotic stress. These genes are responsible either for signaling or activation of some regulatory pathways, or encode some enzymes which involves in the synthesis of structural and functional protectants [31]. HSP are produced by plants in stress conditions especially in response to high temperature. At high temperatures many proteins and enzymes lose their conformation and become dysfunctional. HSPs are shown to guard 75% of the total soluble proteins in vitro from heat denaturation. Under stress conditions, HSPs perform like molecular chaperons and maintain the protein structure. There are five conserved families of HSPs i.e., HSP100, HSP90, HSP70, HSP60 and sHSP. Among them sHSPs (small heat-shock proteins) is widespread in plants. HSPs molecular sizes ranging 15 to 30 kDa. All sHSPs are encoded by the nuclear genes. The defensive effects of HSPs can be endorsed to the chaperones network which acts coordinately [32]. Across the cell membrane following osmotic or hydrostatic pressure gradients the water regulation is regulated by a class of proteins known as aquaporins. Aquaporins are biologically very important proteins because of their ability to transport water in places where its regulation is physiologically critical. There are two major sub groups of aquaporins i.e., tonoplast intrinsic proteins (TIPs) and the plasma membrane intrinsic proteins (PIPs) [33]. PIP has a major role in Trans cellular water transport and is further divided into PIP1 and PIP2. TIP, on the other hand is involved in the exchange of water between cytosolic and vacuolar compartments and also assist in osmoregulation. Antisense inhibition of PIP aquaporins in some plants resulted in decrease of its stress tolerance ability, which suggests the importance of aquaporins. Over expression of PIP make plant hyper sensitive to stress conditions [34]. Under drought conditions, transgenic plants which over express the PIP showed poor germination and growth of wild plant. For the improvement of abiotic tolerance in plants TIP is more preferable.
Drought Tolerance and the Effect of Silicon
Silicon is an essential element for higher plants because without silicon plants grow in an irregular manner. Irrespective of its importance as an element silicon has also been reported as valuable in alleviating both biotic stresses abiotic stresses [35]. It is because of these properties that silicate fertilizers are good soil adjustment to plant production along with decreased methane gas production in rice farming. Furthermore, if silicon can recover drought tolerance of plants its solicitation may also help alleviate the necessity for irrigation which as a result could decrease salinization of field. Silicon does not corrode or cause pollution so that silicon fertilizers are best choice for emerging ecologically green cultivation [36]. Plant root absorb silicon as silicic acid. This is a stable molecule which is usually present about 0.1 to 0.6 mM concentration in the soil at pHs ranging from 1 to 9. After the uptake of silicic acid it is then moved towards the shoot through xylem. It has been extensively testified that silicon enhances drought tolerance in plants for example in rice, pepper, and cucumber. Silicon also raises heat tolerance by conserving membrane strength. As drought is occasionally escorted by extraordinary temperature usage of silicon can lessen the destruction of together drought and heat [37].
Enhancing Nutritional Phosphorus in Plants
Vesicular-arbuscular mycorrhizal fungi (which help phosphorus intake in plants) are identified to increase drought tolerance. It can arouse the growth and yield of plants grown in well irrigated conditions. The improved size of the mycorrhizal plants in drought is more as compared to non-mycorrhizal plants. Low diffusion of P in normal plants could be improved by the fungus covering a greater volume of the soil result in extending outside the depletion zone which covers the zone of plant roots. This is the explanation for the usual growth of plants with mycorrhizal infection that is observed in fine irrigated soils with low P level. The mycorrhizal fungus reduces the low uptake of P via roots through bringing it directly into the roots from surroundings. But mycorrhizal fungi are pointless when P fertilizers are accessible. Under the natural conditions when soil moistness is little or it experiences cyclic differences, plant growth by mycorrhizal fungi cannot be repeated by adding P. The amplified drought tolerance subsequent from mycorrhizal fungi can help plant growth and shows that mycorrhizal fungi is more important in dry conditions than when water is abundant.
Prospective Implementation
For future concerns, it is necessary to develop such genotypes of plants which have higher tendency to tolerate drought conditions and require less amount of water for the production of biomass. There is a great need of advancements in genomics so that to make transgenic plants more tolerant to the stress conditions. Researchers are working on to see the effects of various substances such as osmoprotectants, minerals, acids, and growth regulators towards the induction of drought resistant plants. Several methods including mass screening along with breeding are being used to look for the genetic variation to select those plants which are resistant to droughts and to introduce the same genes to the other plants for better agronomic traits [38]. Many different strategies are being developed to induce the early onset of drought escape, modify the plant morphology to help plants tolerate drought conditions, and to introduce drought resistant characteristics for a greater grain yield along with the water retention system [39]. Although it is difficult to identify and clone drought tolerant genes but through scientific efforts it has been observed that C4 path-ways possess high photosynthetic rates with better water retention and high amount of biomass formation. It happens especially when the internal carbon dioxide concentration in the leaves is low and the temperature in the environment is very high. In case of C3 plants, researchers are looking into ways to over produce PEP-Carboxylase enzyme to enhance the photosynthesis for a better grain yield even in drought conditions. It has already been observed that the transgenic rice crops produce satisfying amount of PEP-Carboxylase and hence tolerate drought conditions by having a very low reduction in the tare of photosynthesis. Through this, scientists now believe that the over production of this enzyme could help in enhancing photosynthesis in plants [40].
Percentage Decrease in the Grain Yield of Plants
During drought period, there are many metabolic changes which occur in plants. Many responses include the shutting of stomata, K+ and Na+ accumulation and damage of plant cell membrane, varied production of different enzymes including those enzymes which are directly involved in the formation of ATP, and the production of phytohormones. The plant phytohormones and other special substances of great attention include auxins, salicylic acids, cytokinins, abscisic acid, and gibberellins (Figure 1). It has been observed that the severe conditions of drought are, the more plant grain yield decreases due to low the accumulation of K+ and Na+ in plant cells which are then pumped either in the vacuole or are removed out of the cells.

Figure 1: Decrease in percentage grain yield in rice and wheat [41-44].
Role of Various Genes, Proteins, and Amino Acids
It describes many genes including Beta, Cox, TPSP, ERA1, CDSP32, and p5csf129a which help in overcoming drought conditions and in osmoregulation. The mechanisms and usefulness has been describes therewith. Besides many genes which produce enzymatic proteins for reduction in ROC’s there are some useful biomolecules such as proline which helps in striving against drought (Table 1). Proline is an amino acid released in plants especially during drought stress. A low water potential is maintained with the help of proline through osmoregulation. Levels of proline in cells also have a wide effect on gaseous exchange, activities of enzymes involved as anti-oxidants, and the relation between water and plants. It is the gene p5csf129a which codes for the special kind of enzyme that helps in producing proline in abundant amount when needed. Presence of abundant proline endogenously in cell helps in maintenance of turgor pressure.
| Plant Genes/Proteins | Host Plants Mechanism | Plant Source |
|---|---|---|
| Beta Gene | Cotton Crops | Accumulates GB to reduce drought tolerance in cotton crops. |
| Wheat Crops | Accumulates GB to reduce drought tolerance in wheat plants. | |
| Cox Gene | Arabidopsis Plant | Accumulates GB to reduce drought Tolerance in Arabidopsis Plants. |
| Tobacco Plant | Accumulates GB to reduce drought Tolerance in Tobacco Plants. | |
| TPSP | Rice Crops | Protection of Plants |
| Rice Crops | Effective Photosynthesis | |
| ERA 1 Farnesyl Transferase | Canola Plant | Farnesyl Transferase downregulates and as a result ABA produces to prevent drought. |
| Canola Plant | Reduced Stomatal pore conductance | |
| p5csf129a Gene | Tobacco Plant | Increased Production of Proline amino acid. |
| Various Plants | Increased turgor pressure in plants. | |
| CDSP 32 | Potato Plant | Protection of the Chloroplast from ROC’s |
| Potato Plant | Prevention of Oxidative Damage. | |
| Other genes | Maize | Improved Photosynthetic machinery |
| Tobacco | Increase in Stomatal pore conductance |
Note: TPSP: Trehalose-6-Phosphate Synthase Phosphatase; CDSP: Chloroplastic drought-induced stress protein; GB: Glycine betaine; ROC: Reactive Oxidation Species[45-50]
Table 1. Role of various genes and proteins in different plants.
Drought Resistance and Improved Yield in Palm Oil
We need advanced techniques of genotyping and genome sequencing to prevent usage of large agriculture lands but using small lands having crops with high yields and more resistance towards drought and diseases (Figure 2). the genome sequencing of Oil Palm increased the yield to 4 tons per Ha as compared to the other oil crops which had average of 0.5 tons per Ha and occupied large land area. It is due to advancements of crop genotyping that plants are now more tolerant towards drought which leads to greater yield. Oil palm not only occupied less area but also helped in conservation of biodiversity.

Figure 2: Effect of genome sequencing on palm oil towards greater yield and high resistance from drought, diseases, and salinity [51].
Of many environmental constraints which crop plants have to bear; Soil Drought is one, which would keep increasing towards severity in coming years. Drought is a condition of reduced or very low availability of water to the plants for a prolonged period of time which disturbs growth, development, water relations, and efficiency of many terrestrial plants. To overcome drought conditions, plants acclimatize themselves to different biochemical, physiological, morphological changes which are quite sophisticated. So far many drought tolerance plants have been developed, despite of the limitations of conventional breeding. Cuticle development, controlled stomatal opening, genetic control and other biotic and abiotic factors were controlled to produce the drought tolerance plants. During evolution, certain processes were seen which have evolved plants that now they can be tolerant to prone drought. C photosynthesis and development of crasulaccean acid metabolism have shown the increase in water efficiency. Deep rooting and further cuticle deposition, breeding, genetically modified crops to conserve water. Water is required mainly by plant for metabolic activity. Physiological tests have shown better plants to increase the progress for the better developed plants. For the improvement of crop, the biology, climate, physiology and chemistry of the plant should be better understood for the development of drought tolerance plants.
We’d like to pay thanks to Dr. Muhammad Khurshid for helping us with all our enquiries relevant to the topics under discussion and for his commendably supportive comments which greatly improved this manuscript. We’d also thank all the reviewers of this review paper, for their great comprehension, time, and endeavors they’ve put in reviewing this manuscript.