Research & Reviews: Research Journal of Biology
ISSN: 2322-0066
ISSN: 2322-0066
Guo Ming Shen*
Department of Agriculture and Bioengineering, Heze University, Heze, China
Received: 28-Aug-2023, Manuscript No. JOB-23-112624; Editor assigned: 31-Aug-2023, PreQC No. JOB-23-112624 (PQ); Reviewed: 14-Sep-2023, QC No. JOB-23-112624; Revised: 21-Sep-2023, Manuscript No. JOB-23-112624 (R); Published: 28-Sep-2023, DOI: 10.4172/2322-0066.11.3.005
Citation: Shen GM. Effects of Epibrassinolide on Wheat Seedling Growth and Antioxidant Properties Under Cd2+ Stress. RRJ Biol. 2023;11:005
Copyright: © 2023 Shen GM. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Visit for more related articles at Research & Reviews: Research Journal of Biology
This study investigates the physiological changes of two varieties wheat (Triticum aestivum L.) seedlings namely LX77 and JM22 by spraying different concentrations of Epibrassinolide (EBR) under Cd2+ stress using a hydroculture. Results showed that after foliar application of different concentrations of EBR, seedling growth properties, chlorophyll contents, root vigor, osmotic regulators, Malondialdehyde (MDA) and H2O2 contents, relative conductivity, antioxidant enzyme activities were changed in both LX77 and JM22, and the optimal exogenous EBR concentration is 0.01 or 0.1 mg·L-1. The survey conducted by Chinese Agriculture Ministry showed, the sewage irrigation area is about 1.4 million hm2, which suffered heavy metal pollution of the land area accounted for 64.8%, Mercury (Hg) and Cd are the largest pollutant.
Wheat; Epibrassinolide; Cadmium stress; Seedling growth; Antioxidant properties
EBR: Epibrassinolide; MDA: Malondialdehyde; POD: Peroxidase; CAT: Catalase; AsA: Ascorbate; GSH: Glutathione; LX77: Liangxing 77; JM22: Jimai22
With the rapid development of industry and agriculture, the excessive development of mineral resources and irrational use of chemical fertilizers, Cadmium (Cd) has become one of the major heavy metal pollutants to environment and hurt human beings health through food chain [1,2]. China's annual heavy metal pollution caused by more than 10 million ton food production, contaminated by heavy metal food up to 12 million tons, the total economic loss of at least 20 billion yuan, the reduction of Chinese grain production due to heavy metal pollution exceeds 10 million tons every year, grains polluted by heavy metals as many as 12 million tons, leaded at least 20 billion yuan economic losses [3].
For Cd is a non-essential element of plant growth, it has strong chemical activity, mobility and biological toxicity, and it is easily absorbed and accumulated by plant roots and is easily transported to the ground [4-6]. Cd contamination causes a series of damage to plant growth and development, which reduces plant root length, plant height and biomass, and inhibits photosynthesis of plants [7,8]. Excessive Cd not only affects the growth and development of plants, but also enters the human body from the food chain through edible parts such as grains, fruits, and vegetables. When Cd enters the human body, it will not decompose and accumulate in the human body, causing teratogenicity, carcinogenesis, mutation, etc. [9,10]. Because China has no more land, it is difficult to reduce the loss of crop growth caused by Cd by using large-scale land fallow to repair pollution. Therefore, choosing a suitable method to reduce the harm of Cd to crop growth is a way to solve the problem.
Brassinolide, as a phytohormone commonly known as Brassinosteroid (BR), is a natural compound found in plants, and more than 70 species of BRs have been discovered [11-13]. Studies have shown that BRs can promote plant growth by participating in plant cell division and cell elongation, photosynthesis and other physiological processes, effectively reducing the damage of biotic and abiotic stress [14-18]. 2,4-Epibrassinolide (EBR) is a synthetic high-activity BR analogue that enhances the growth and development of a variety of crops under a variety of stresses [19-21]. Wheat ( Triticum aestivumlinn L.) is the third largest food crop in the world and China. It is also the main food crop in northern China. Its stable and high yield has important strategic significance for the development of the national economy. Therefore, it is important to clarify the effect of Cd on wheat growth and explore ways to alleviate cadmium stress on the safe and high quality production of wheat in China. Studies have shown that a certain concentration of EBR can alleviate the effects of salt or alkali, Hg and other abiotic stresses on wheat growth and development [22,23]. In view of this, we wanted to explore whether EBR also has the similar biological function, under Cd stress in wheat. The Cd-tolerant Liangxing 77 (LX77) and the more resistant Jimai 22 (JM22) screened in our previous experiments were used as materials to study the effects of EBR on wheat seedling growth and antioxidant properties under Cd stress. The effects of EBR on the wheat under Cd stress were explored to provide a theoretical basis for the use of EBR to alleviate the damage of Cd on wheat, and to lay a foundation for further study on the mechanism of EBR regulating Cd toxicity in wheat.
Plants materials and reagents
Two varieties of wheat, namely Liangxing 77 (LX77) and Jimai 22 (JM22) were purchased from Heze Seed Company (Heze, China). EBR (CAS No.: 78821-43-9) was purchased from Merck KGaA (Darmstadt, Germany) other chemicals and reagents were of analytical grade and purchased from Shenggong Chemicals Co. Ltd. (Shanghai, China).
Sampling
The evenly full wheat seeds were selected and disinfected with 10% sodium hypochlorite for 10 min. The distilled water was rinsed repeatedly at least for 3 times, and then soaked in a constant temperature incubator at 25˚C for 12 h, germinated in a moistened filter paper with half-strength (1/2) Hoagland nutrient solution at 30˚C for 2 d. Pick out the same white wheat seeds, planted them in the germination box, and pour the 1/2 Hoagland nutrient solution every 2 d after the cotyledons are flattened. Cultivate to the three-leaf one heart stage. Wheat seedlings with healthy and uniform growth were incubated in 1/2 Hoagland nutrient solution for 3 days.
The concentration of CdCl2 was 50 mg·L-1, and the concentration of EBR was 0, 0.001, 0.01, 0.1 and 1.0 mg·L-1 were used in this experiment. A CdCl2 solution with a concentration of 50 mg·L−1 was prepared as a hydroponic solution with 1/2 Hoagland nutrient solution, and the wheat seedlings with uniform growth were taken for treatment: 1) CK, hydroponic solution was 1/2 Hoagland nutrient solution (excluding CdCl2) ), foliar spray distilled water; 2) hydroponic solution is a solution of 50 mg·L-1 CdCl2 prepared with 1/2 Hoagland nutrient solution (the same below), foliar spray distilled water; 3) foliar spray 0.001 to 1.0 mg·L-1 EBR. EBR (or distilled water) should be sprayed on the surface at 16:00 every day. It is better to use both sides of the blade to be wet and no liquid drops. Replace the hydroponic solution every 2 d. 20 seedlings per bottle, repeated 3 times, cultured in an artificial climate chamber (JPGX-1000C, Shanghai Jinwen Instrument Equipment Co., LTD, Shanghai, China) with a temperature of 20/25˚C, an illumination of 4000 to 4500 Lx, and a light/dark cycle of 14 h/10 h. The 10 d samples were cultured for the determination of growth indicators and physiological indicators.
Determination of growth indicators and Cd contents
After 10 days of treatment, 10 wheat seedlings were randomly selected for each iteration. After the plant height was measured, cut from the roots and shoots, placed in an oven at 110˚C for 5 min, then dried at 80˚C to constant weight, using an electronic balance (BSA124S-CW, Sartorius AG, Goettinge, Germany) to weight the shoots and the roots Dry Mass (DM) and averaged. Cd contents were measured using Atomic-Absorption Spectroscopy (AAS, Perkin Elmer, Inc., Waltham, MA, USA) after incubation with HNO3-HClO4 (4:1) at 350˚C for 20 h.
Determination of physiological and biochemical indicators
After 10 days of treatment, the same node leaves of wheat seedlings were taken, rinsed with distilled water for 5 times, cut and mixed, and used for physiological index determination, and each treatment was repeated triplicate. The chlorophyll contents were determined by extraction method and determined by RG-5000T spectrophotometer (Tianjin Nuore Xinda Technology Co., LTD, Tianjin, China). The root activity was determined by TTC method, the soluble protein content was determined by Coomassie brilliant blue method, and the soluble sugar content was determined by the fluorenone method [24,25]. Determination of SOD, POD and CAT activities and MDA, Hydrogen peroxide (H2O2), Ascorbic Acid (AsA) and reduced Glutathione (GSH) contents was described by Shen, et al. [26]
Statistical analysis
ANOVA Duncan's multiple range test was used for statistical analysis of each group. All statistical analyses were performed using SAS 9.4 (SAS Institute Inc.,). A probability p<0.05 value is considered significant. The values expressed for each treatment are the means of three experimental repetitions.
Effect of EBR on plant shoot or root length and root numbers of wheat seedlings under Cd stress
The shoot and root length of the two varieties were decreased significantly after treatment with 50 mg·L-1 Cd2+ compared with the control, with a decrease of 17.98% and 12.15%, respectively, while in root, with a decrease of 62.93% and 39.49%, respectively. The shoot and root length of both varieties were increased first and then decreased after foliar application of different concentrations of EBR. The effect of EBR on LX77 is 0.1 mg·L-1 treatment reaching the maximum, which was 25.37% higher than that of Cd2+ alone, while in root the concentration is 0.1 mg·L-1 and share in 50.86% increase. The shoot and root length of JM22 reached the maximum when the EBR concentration is 0.01 mg·L-1, which was 19.85% and 48.09% higher than that of Cd2+alone, respectively. EBR treatment with 0.01 mg·L-1 and 0.1 mg·L-1 concentrations can obviously promote the growth of shoot and root, other concentrations were no significant difference compared to the control under Cd stress, indicating that spraying a certain concentration of EBR on leaf surface could promote the growth of wheat seedlings. The root number of the two varieties increased by 7.71% and 33.27%, respectively, after treatment with 50 mg·L-1 Cd2+. Compared with Cd2+ treatment alone, the number of roots of the two varieties were decreased first and then increased after foliar application of different concentrations of EBR, the optimal concentration of rooting inhibition is 0.001 mg·L-1 rather than 0.01 mg·L-1 of external EBR supply. EBR treatment showed no significant difference in root reduction, indicating that EBR was ineffective in this biological process. We speculate that one of the mechanisms of resistance of JM22 to Cd was realized by increasing root segregation (Figures 1A-1F).

Figure 1: Effect of EBR on plant height and root numbers of wheat seedlings under Cd stress. The data are means ± SD; bars with different letter(s) are significantly different at p ≤ 0.05. 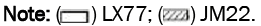
Effect of EBR on biomass and Cd contents of wheat seedling under Cd stress
With the treatment of 50 mg·L-1 Cd2+, the biomass of two varieties were decreased significantly both in shoot and root, with a decrease of 17.71% and 15.18% in shoots, and 24.34% and 20.41% in roots respectively. Foliar application of different concentrations of EBR can increase the biomass, when EBR concentration is 0.1 mg·L-1, biomass accumulation can reach the level of Cd free, indicating suitable EBR treatment could reverse the biomass loss caused by Cd toxicity. It is clear that plants under Cd stress accumulate more Cd in the roots and then transfer it to the shoots via xylem transport [26]. The Cd contents of shoot of both varieties were decreased first and then increased after foliar application of different concentrations of EBR, and the opposite occurs in the roots. The accumulation of Cd in shoots and roots of all exogenous EBR treatments reached statistically significant differences, indicating that exogenous EBR treatment could promote Cd accumulation in plants. LX77 accumulated more Cd than JM22 both in shoots and roots, indicating that LX77 is a tolerance type wheat of Cd (Figures 2A-2H).

Figure 2: Effect of EBR on biomass and Cd contents of wheat seedling under Cd stress. The data are means ± SD; bars with different letter(s) are significantly different at p<0.05.
Effect of EBR on chlorophyll contents, root vigor and osmotic regulators of wheat seedlings under Cd stress
The chlorophyll contents of the LX77 and JM22 did not show statistically significant differences after treatment with 50 mg·L-1 Cd2+ compared with the control. Compared with Cd2+ treatment alone, the chlorophyll contents of LX77 were basically unchanged after foliar application of different concentrations of EBR, only the EBR concentration were reached 0.1 or 1 mg·L-1, the chlorophyll contents were increased significantly by 68.89% and 71.11%, respectively. It is indicated that EBR of 0.1 or 1 mg·L-1 can promote the synthesis of chlorophyll in LX77 leaves which may be one of its adaptive strategy under Cd stress. Interestingly, the contents of chlorophyll in JM22 were decreased instead when external EBR were reached the concentration of 1 mg·L-1, indicating high-dose EBR has the opposite effect. Cd seriously inhibited the root vigor of LX77 by 41.89% compared to control, but had no effect on JM22, which may be one of the mechanisms of JM22 anti-Cd. The root vigor of LX77 were decreased first and then increased after foliar application of different concentrations of EBR compared to control. When the EBR concentration was 0.1 mg·L-1 and 1 mg·L-1, the root vigor increased by 41.86% and 62.79%, respectively. Compared with Cd2+ treatment alone, the root vigor of JM22 changed little after exogenous EBR spraying was 0.001 mg·L-1, when it reached 0.01, 0.1 and 1 mg·L-1, the root vigor of JM22 were increased by 86.67%, 60.00% and 40.00%, respectively. It indicated that spraying exogenous EBR on leaves could improve root vigor in both LX77 and JM22. Treatment with 50 mg·L-1 Cd2+ alone, the soluble protein contents of two varieties did not statistically significant differences between LX77 and JM22 compared to control. Compared with Cd2+ treatment alone, the soluble protein contents of LX77 was increased first and then decreased after foliar application of different concentrations of EBR, and reached the maximum when the EBR concentration was 0.1 mg·L-1, and it was increased by 37.21%, indicating the exogenous EBR could promote soluble proteins biosynthesis. There was no significant differences between the soluble protein contents of JM22 at different EBR concentration treatment, indicating the effect of exogenous EBR on JM22 soluble proteins was not obvious. Compared with the control, the soluble sugar contents in both varieties were increased at statistically significant differences level after treatment with 50 mg·L-1 Cd2+, indicating Cd treatment will promote the synthesis of soluble sugars in them. After spraying different concentrations of EBR, the soluble sugars contents of LX77 were decreased first and then increased and remained at a high level, but there was no significant differences compared with Cd2+ treatment alone. Foliar application of different concentrations of EBR to JM22 enhance soluble sugars accumulation, especially with 0.1 mg·L-1 EBR treatment was increased by 89.47% indicating that soluble sugars may play a critical role in JM22 anti Cd toxicity (Figures 3A-3H).
Effect of EBR on AsA, GSH, MDA or H2O2 contents and relative conductivity of wheat seedlings under Cd stress

Figure 3: Effect of EBR on chlorophyll contents, root vigor, and osmotic regulators of wheat seedlings under Cd stress. The data are means ± SD; bars with different letter(s) are significantly different at p<0.05.
Compared with the control, the AsA and GSH contents of the two varieties were decreased significantly after treatment with 50 mg·L-1 Cd2+ alone, with a decrease of 33.10% and 19.75% of AsA or 42.11% and 23.81% of GSH, respectively. Compared with Cd2+ treatment alone, the AsA and GSH contents of the two varieties were increased after foliar application of different concentrations of EBR, and the concentration of 0.1 mg·L-1 was the most effective one, these results indicated that exogenous EBR could promote antioxidants biosynthesis. The contents of MDA and H2O2 were opposite to former ones. Foliar application of exogenous EBR could reduce these harm substances accumulation
The relative conductivity of LX77 JM22 did not show statistically significant differences after treatment with 50 mg·L-1 Cd2+ compared with the control. After treatment with different concentrations of EBR, the relative conductivity of the two varieties were decreased first and then increased, and the lowest when the EBR concentration was 0.01 mg·L-1, the decrease was 62.09% and 53.38%, respectively, indicating that the EBR treatment can be reduced relative conductivity of wheat seedlings at a certain concentration under Cd2+ stress (Figures 4A-4J).

Figure 4: Effect of EBR on AsA, GSH, MDA or H2O2 contents and relative conductivity of wheat seedlings under Cd stress. The data are means ± SD; bars with different letter(s) are significantly different at p<0.05.
Effect of EBR on antioxidant enzyme activities of wheat seedlings under Cd stress
The SOD activities of LX77 were decreased significantly after treatment with 50 mg·L-1 Cd2+, which was decreased by 13.74%, and there was no statistically significant differences in JM22. Compared with Cd2+ treatment alone, the SOD activities of the two cultivars were increased first and then decreased after foliar application of different concentrations of EBR. Exogenous EBR can enhance SOD activities in both LX77 and JM22 indicating that EBR may play a useful role for scavenge Reactive Oxygen Species (ROS). The optimal EBR concentrations are between 0.01 and 0.1 mg·L-1.
Compared with the control, the POD activities of LX77 were increased after treatment with 50 mg·L-1 Cd2+, with an increase of 35.38%, while there was no statistically significant differences in JM22. The POD activities of LX77 were decreased first and then increased after foliar application of different concentrations of EBR, and reached the maximum when EBR concentration was 1 mg·L-1 just as control. The POD activities of JM22 were decreased first, then increased, then decreased and then increased, but the EBR concentration did not change significantly between 0.001 to 1 mg·L-1 (Figures 5A-5F).
Those results indicating that EBR could not stimulate the POD activities of wheat seedlings under Cd2+ treatment. The CAT activities of LX77 were decreased while they were increased in JM22 after treatment with 50 mg·L-1 Cd2+. Compared with Cd2+ treatment alone, after spraying EBR with different concentrations, the CAT activities of LX77 were decreased and then increased. When the EBR concentration was 1 mg·L-1 reaching its peak, but there were no statistically significant differences compared to control. The CAT activities in JM22 were increased and then decreased, and reached the maximum when the concentration of EBR was 0.01 mg•L-1, which was 13.23% higher than that of control. It is indicated that the CAT activities of different wheat cultivars had different EBR concentration responses under Cd2+ stress. A certain concentration of EBR could increase the CAT activities in both LX77 and JM22.
As a non-essential element of plants, Cd is one of the most toxic substances in heavy metal toxicity. Cd stress can induce plants to produce Reactive Oxygen Species (ROS), leading to cell membrane peroxidation, causing disturbance of plant physiological processes and inhibiting plant growth [27]. Bioaccumulation comprehensively reflects the degree of physiological damage to plants, and is a direct manifestation of plants' response to stress [28]. Studies have shown that a certain concentration of Cd2+ inhibits the growth of wheat seedlings and reduces their biomass [29-32]. The results of this study showed that 50 mg·L-1 Cd2+ treatment inhibited the growth of two wheat seedlings, and the plant height, shoot and root dry weight of the two wheat seedlings decreased, which is consistent with the results of Chang, et al.[31]. Foliar application of a certain concentration of EBR can promote the growth of wheat seedlings, increase the plant heights and biomass of wheat seedlings, and the effective concentration of EBR is 0.1 mg·L-1, our results are consistent with the results as regards concerning a certain concentration of EBR can relieve cadmium on soybean seedling growth, and the different concentrations of EBR required may be due to different experimental materials [33]. It has been reported that Cd can induce changes in root morphology and number, in this investigation, treatment with Cd2+ alone increased the number of roots of the two varieties of wheat, which may be due to the ability of wheat to increase its absorption capacity to accommodate Cd2+ stress, while EBR treatment showed little effects on root number reduction, indicating that EBR was ineffective in this biological process. Our previous study has confirmed that exogenous ABA can promote Cd accumulation in roots and reduce Cd accumulation in leaves, this study found that the effect of EBR on Cd accumulation was similar to that of ABA [34,35]. Photosynthesis is an important basic physiological activity of plants. Photosynthetic pigments are important pigment molecules for photosynthesis, and chlorophyll is the most important one [36,37]. Studies have shown that excessive cadmium stress leads to plant chloroplast and pigment disintegration [38,39]. The results of this study showed that there was no significant change in chlorophyll content of the two wheat cultivars after treatment with 50 mg·L-1 Cd2+, indicating that the leaves of two varieties of wheat seedlings chlorophyll synthesis did not change much, probably because 50 mg·L-1 Cd2+ could not inhibit or promote the chlorophyll synthesis of these two varieties of wheat. Foliar application of EBR of 0.01 or 0.01 mg·L-1 had little effect on the chlorophyll content of LX77 and JM22 seedlings, but the EBR of 0.1 or 1.0 mg·L-1 was sprayed on the foliage, the chlorophyll content of LX77 seedlings increased significantly. While the concentration of EBR is 1.0 mg·L-1 which significantly reduced the chlorophyll contents of JM22 seedlings, indicating that different wheat varieties responded differently to high concentration EBR. However, the leaf chlorophyll contents of JM22 are always higher than LX77, indicating that JM22 may be have an efficient mechanism of chlorophyll synthesis.
The vitality level and growth of roots directly affect the nutrient status and yield of above-ground parts, and root vigor is one of the key indicators of roots to characterize the roots of plants [40,41]. The results showed that the root vigor of different varieties of wheat seedlings had different responses to 50 mg·L-1 Cd2+ treatment. The root vigor of LX77 seedlings decreased significantly, and the root vigor of JM22 seedlings did not change significantly. It indicated that the roots of JM22 wheat seedlings were more resistant to Cd2+ stress. After spraying a certain concentration of EBR, the root vigor of LX77 seedlings decreased first and then increased, and increased significantly when the EBR concentrations were 0.1 or 1 mg·L-1, the root vigor of JM22 with the similar effects, indicating that the foliar application of EBR can improve the root vigor, similarly biological functions of GA and SADH in grape shoot vigor [42]. Osmotic regulation is a physiological response of plants to adapt to stress, and soluble proteins and sugars are important osmotic adjustment substances. This study found that 50 mg·L-1 Cd2+ treatment alone inhibited the synthesis of soluble protein in LX77 seedlings, and promoted the synthesis of soluble protein in JM22 seedlings, indicating that the sensitivity of different wheat varieties to Cd2+ stress are different. After spraying EBR with different concentrations, the soluble protein content of the two wheat cultivars increased first and then decreased, and reached the maximum when the EBR concentration was 0.1 mg·L-1, indicating that a certain concentration of EBR could promote soluble protein synthesis of wheat seedlings under Cd stress. This study showed that 50 mg·L-1 Cd2+ treatment promoted the synthesis of soluble sugars in two varieties of wheat seedlings, which increased the soluble sugar content of both varieties. Foliar application of different concentrations of EBR could not increase the soluble sugar contents of LX77 seedlings, but promoted the increase of soluble sugar content of JM22. It is suggested that the soluble sugar synthesis in different varieties of wheat seedlings has different sensitivity to EBR. AsA and GSH are the main antioxidants in plants, which can eliminate excess ROS produced during cell metabolism and reduce the damage caused by membrane lipid peroxidation [43]. It also plays an important role in free radical scavenging reactions in plants [44]. The results of this study showed that the content of AsA and GSH in the two wheat cultivars decreased significantly after treatment with 50 mg·L-1 Cd2+ alone, and the decrease level of JM22 was smaller than LX77, indicating that the resistance of JM22 to Cd2+ stress was stronger than that of LX77. After spraying EBR at different concentrations, the contents of AsA and GSH in the two wheat cultivars increased first and then decreased, indicating that the EBR could increase the AsA and GSH contents in a range of concentrations of EBR to the two wheat cultivars under Cd2+ stress. Excessive Cd2+ will directly destroy the cell membrane, causing the loss of intracellular substances and the peroxidation of cell membrane lipids, leading to the accumulation of ROS. H2O2 is the main form of ROS, and MDA is the product of membrane lipid peroxidation. Therefore, H2O2 and The level of MDA reflects to some extent the degree of damage to plant cells shown that cadmium stress leads to an increase in MDA contents in leaves of Solanum nigrum L., and a certain concentration of EBR can reduce the MDA contents of Solanum nigrum leaves under cadmium stress [45-47]. The results of this study showed that the H2O2 and MDA contents of the seedlings of LX77 seedlings were increased significantly after treatment with 50 mg·L-1 Cd2+ alone, indicating that Cd2+ caused the cell membrane lipid peroxidation of LX77 seedlings. The MDA contents of JM22 seedlings did not change significantly, and the H2O2 contents increased significantly but the increasing rate was less than that of LX77. The relative conductivity of roots did not change much in both LX77 and JM22, indicating that 50 mg·L-1 Cd2+ affected the relative conductivity of roots in two varieties are limited. Compared with Cd2+ treatment alone, the MDA and H2O2 contents and relative conductivity of the two wheat cultivars decreased to some extent after foliar application of different concentrations of EBR. The EBR concentrations were 0.01 or 0.1 mg·L-1 are the best. Enzymes and non-enzymatic systems in plants resist oxidative stress. Studies have found that the synergistic effect of enzymatic antioxidant system and non-enzymatic antioxidant system can effectively alleviate the toxicity of ROS accumulation induced by stress in wheat [48].
Antioxidant enzymes SOD, POD and CAT are important protective enzymes for scavenging ROS in plants. They can synergistically and effectively eliminate ROS and reduce the damage of plant cells. Study has shown that foliar application of 0.5 mg·L-1 EBR can increase the SOD, POD and CAT activities of soybean seedlings under cadmium stress. This study found that the SOD activities of LX77 seedlings were decreased significantly after treatment with 50 mg·L-1 Cd2+ alone, while the SOD activities of wheat seedlings of JM22 were increased a little, indicating that Cd2+ stress inhibited the LX77 seedlings heavily. Compared with Cd2+ treatment alone, the SOD activities of both wheat cultivars increased first and then decreased after foliar application of different concentrations of EBR. Indicating that foliar application of a certain concentration of EBR could increase the SOD activity of wheat seedlings under Cd stress. After treatment with 50 mg·L-1 Cd2+ alone, the POD activities of both wheat cultivars increased, and the leaves were sprayed with different concentrations of EBR, the POD activities of wheat cultivars did not increase significantly, indicating that EBR could not stimulate the POD activities of wheat seedlings under Cd2+ stress. After treatment with 50 mg·L-1 Cd2+ alone, the CAT activities of LX77 seedlings were increased while the CAT activities of JM22 seedlings were decreased, indicating that the CAT activities of different varieties of wheat varied.
In summary, under Cd2+ stress, wheat seedlings were oxidized and inhibited plant growth. Foliar application of 0.1 mg·L-1 EBR increased the antioxidant enzyme activities and antioxidant contents of leaves, reduced the accumulation of reactive oxygen species, alleviated the damage of oxidative stress on the membrane system, promoted the growth of seedlings, and enhanced the resistance of wheat seedlings under Cd2+ stress. It indicated that one of the possible ways EBR inhibited cadmium-related growth reduction in wheat seedlings is through EBR regulation of antioxidant metabolism. Compared with the treatment of Cd2+ alone, foliar application of a certain concentration of EBR cannot promote CAT activities in both wheat seedlings.
This research was supported by the Natural Science Foundation of Shandong Province (CN) (ZR2022MC203) to SGM, the “5136” talent project of Heze University to SGM and SHQ, and the Doctoral Fund of Heze University (No. XY17BS07) to GCY.
No conflict of interest.
SHQ and GCY participated in the design of experiments, collected the data and drafted the manuscript. SGM participated in the design of experiments and helped write the manuscript. SGM and SHQ coordinated the research and helped to finalize the manuscript. All authors read and approved the final manuscript.
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref] [Google Scholar] [PubMed]
[Crossref]